在地球上,有大量全新和系统发育学上常见的微生物群落在极端环境中生存,这种独特而复杂的生态系统为研究自然微生物群落的结构、功能和进化提供了良好的机会。标记基因研究已经广泛用于研究极端环境微生物群落的生态驱动因素,揭示了大量未培养的微生物群落的多样性,以及在此环境中古菌的优势。而组学研究的新成果展示了微生物群落功能与环境变量之间的密切联系,并可通过基因组表征来发现新的微生物谱系这将极大地扩大了所认知的微生物群落的多样性并改变了系统发育的结构。这些研究极大地推动了我们对地球极端环境中微生物多样性、生态学和进化学的理解,促进了对复杂生态系统中的微生物群落的探索。在本综述中,将总结目前地球主要极端环境中细菌和古菌的多样性、生态学和进化的知识。
主要内容
1. 微生物多样性与主要微生物新谱系
在早期对美国黄石国家公园的陆地地热泉等生物多样性较低的极端环境的原核生物16S rRNA分析改变了过往对地球微生物多样性的理解。随后开展的广泛性微生物基因组分析提供了更加全面的极端环境微生物群落组成和多样性的展示。极端环境由于在全球上离散分布和相对受限的微生物群落结构,因此也特别适合微生物生物地理学研究。仅通过16SrRNA扩增子测序可研究极端微生物群落结构,而宏基因组和单细胞基因组可直接从环境样本中获得接近完整的微生物基因组,提高对其代谢功能和进化的见解。总的来说,对无法培养的微生物的基因组学的研究实质上拓展了生命发育树的广度和深度,特别是古菌领域。
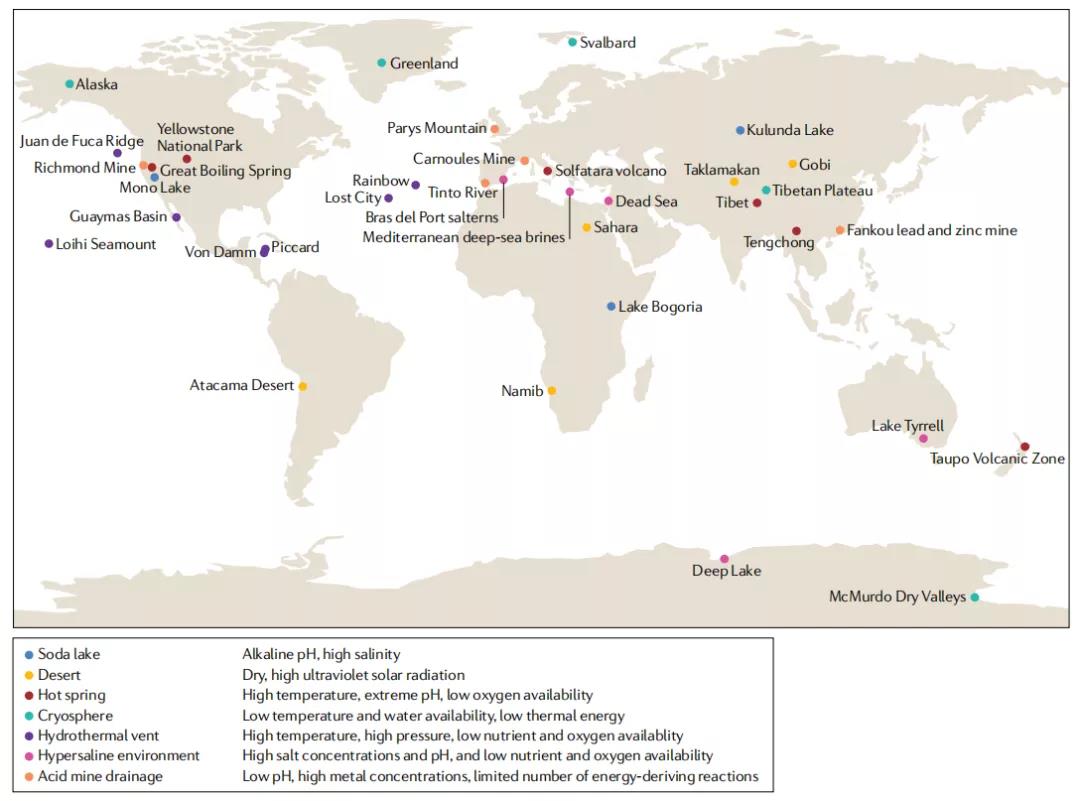
图1 全球的极端环境的分布
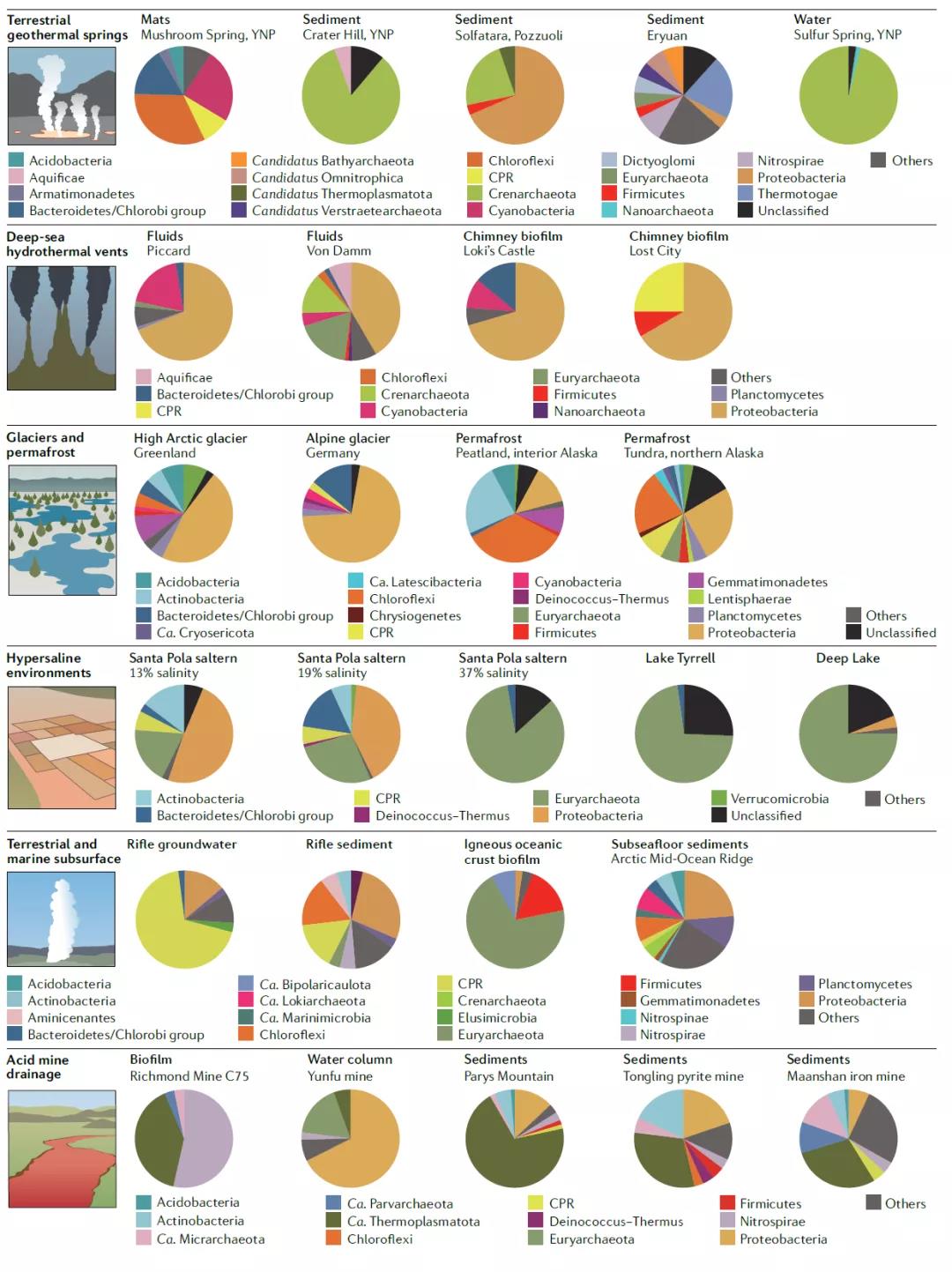
图2 极端环境下的微生物群落组成
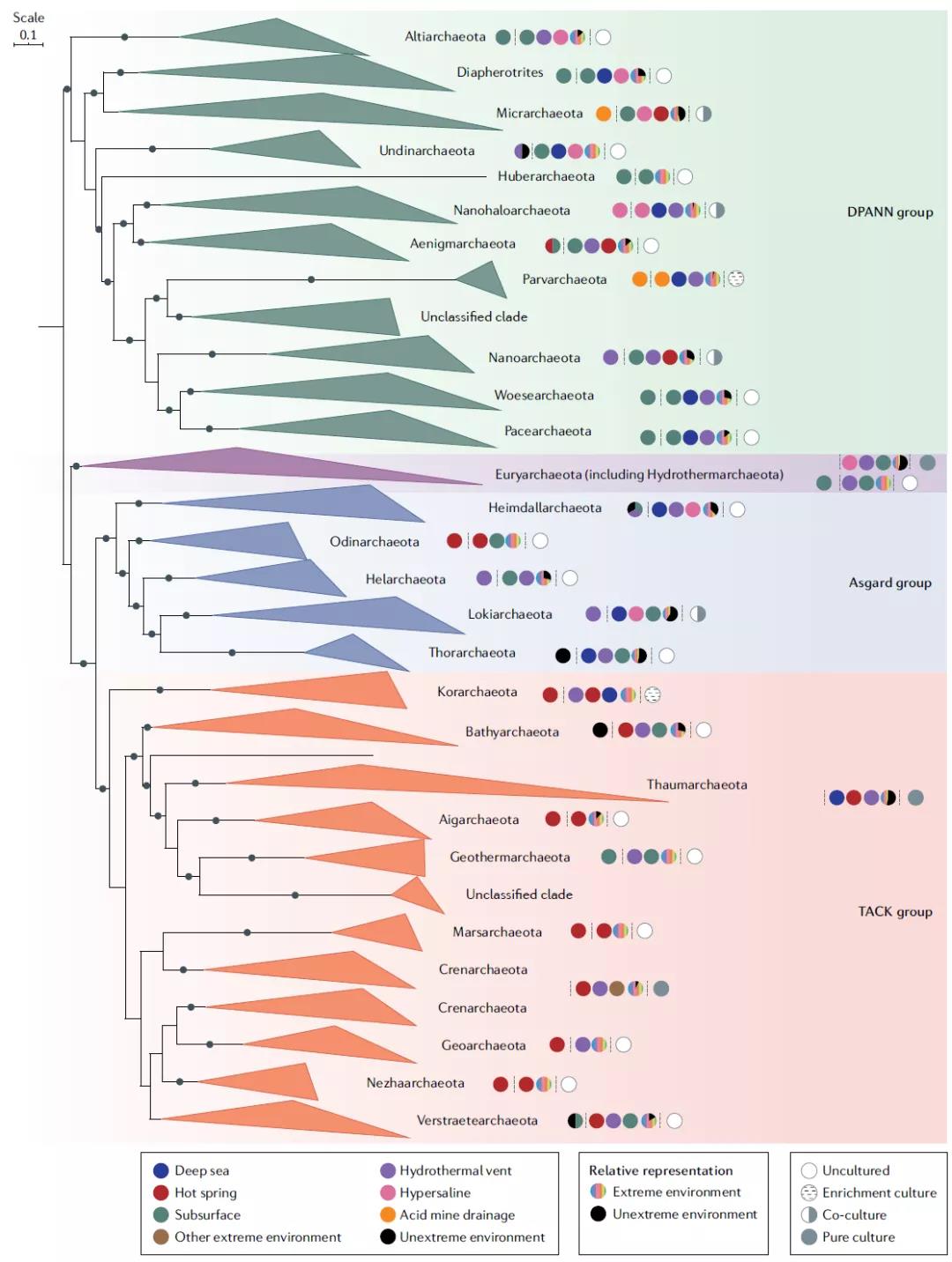
图3 古菌的进化树
2.微生物群落功能与动态
大部分微生物无法培养分离限制了我们对微生物群落的功能在时间与空间上的变化研究。宏组学可直接从环境样本中获得微生物基因组、基因丰度和转录表达等信息,有助于了解微生物群落的功能意义。总的来说,极端环境样本的多种组学数据揭示了极端环境微生物群落参与了能量守恒、碳固定和氮代谢等多方面。同时组学比较分析思路已经应用在探索不同时空尺度的功能变化动力学,可研究地理上独立的微生物群落系统沿着特定环境梯度的变化,可分析群落与环境变量之间的关系,有助于解释在微生物分布中观察到的不同模式。
以AMD为例,组学研究记录了AMD及其相关环境的微生物群落组成,如宏基因组和宏转录组揭示了与AMD中与地球化学密切相关的基因及其转录情况。里士满矿区的AMD位点的嗜酸生物膜蛋白组研究发现丰度最高的细菌的蛋白质受群落组成变化影响最为明显,而丰度较低的细菌的蛋白质表达受非生物因素影响更大。同时,对尾矿不同酸化阶段和AMD生物膜生态演替阶段的微生物群落进行了对比分析,证明了其群落功能与时间变化的密切关系。具体来说,早期和晚期的尾矿群落富含参与低pH和重金属适应的基因,反映了不同的环境胁迫作用。
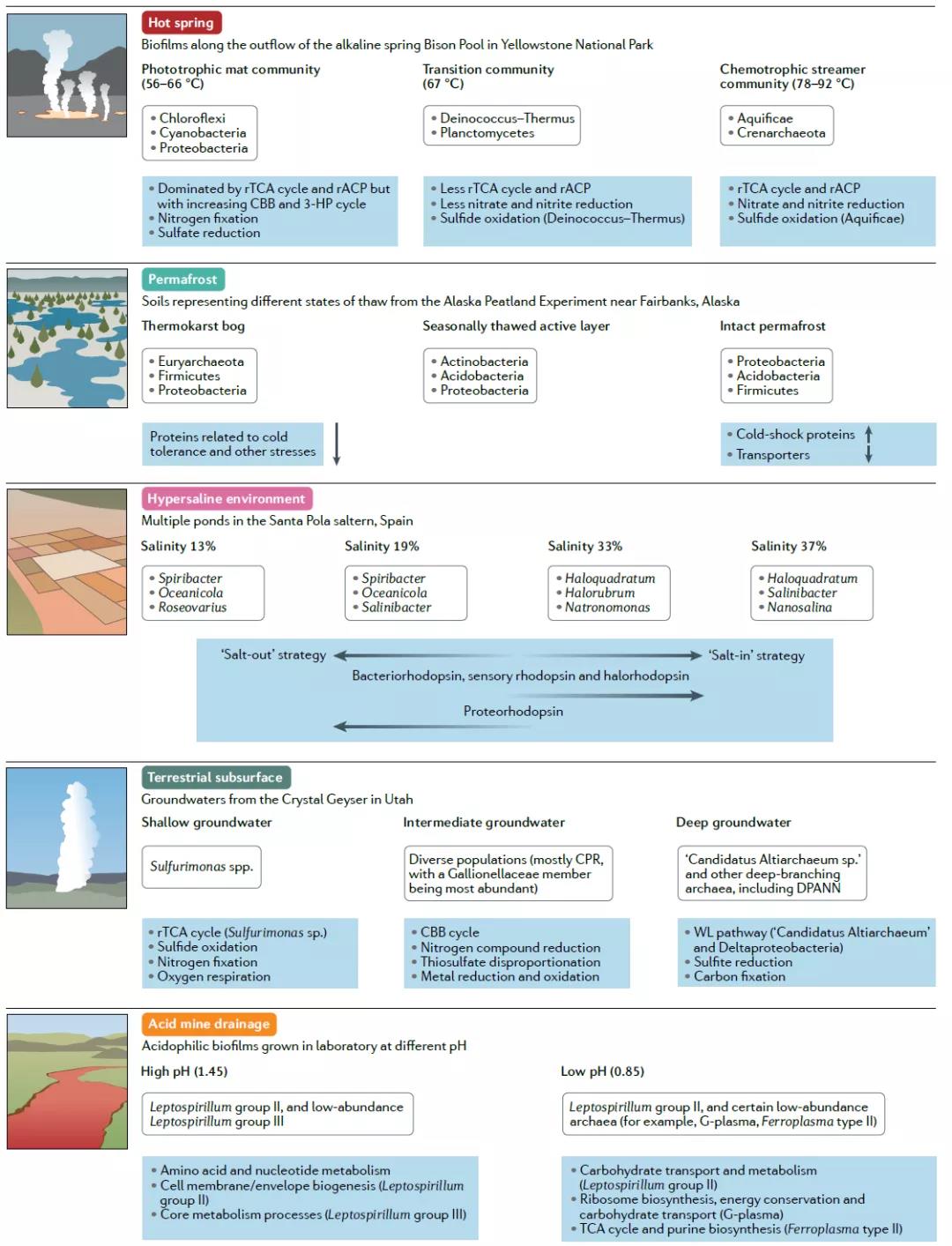
图4 微生物群落在特定环境梯度上的组成和功能转变
3.极端环境中的微生物进化
一直以来,科学家们都在解答微生物是如何适应极端和高度变化的环境,并且在此之中不断进化。这个问题最初是通过模式生物的基因组和生理学研究解决的,如研究微生物基因组的序列和变化模式在自然环境中的存在意义。重组作为进化中非常重要的部分,在嗜盐和嗜热古菌被广泛研究,已有研究表明在盐杆菌属中存在频繁地重组,在富盐菌属中的不同细菌之间甚至发现了长染色体片段交换的高频重组。基因水平转移(HGT)是原核生物基因中基因增加的主要来源,基因含量的变化被认为是微生物适应性进化的主要模式,这情况在嗜盐细菌、古菌和硫化叶菌中尤为普遍。
在努力加深对极端环境的微生物群落的认识过程中的突出问题是这些分析模式是否能应用在限制较少生态系统中,因为没有一个分析模式可以捕捉到所有生态系统的所有特征,最近的研究发现简化模式微生物模式可作为我们对复杂自然微生物群落认知的有效工具。在不断对极端环境中微生物生态和进化的探索将有助于我们对更复杂的自然微生物群落和全球生态系统的理解。
引文:Wen-Sheng Shu, Li-Nan Huang. Microbial diversity in extreme environments. Nature Reviews Microbiology 2021, https://doi.org/10.1038/s41579-021-00648-y