 |
第五章
性别决定与伴性遗传
|
| 第一节 生物的性别决定 | |
| 第二节 外界环境对性别发育的影响 | |
| 第三节 性连锁的发现 | |
| 第四节 染色体学说的直接证据 | |
| 第五节 性相关遗传 | |
| 第六节 人类的性染色体异常 |
|
第五章
性别决定与伴性遗传
|
| 第一节 生物的性别决定 | |
| 第二节 外界环境对性别发育的影响 | |
| 第三节 性连锁的发现 | |
| 第四节 染色体学说的直接证据 | |
| 第五节 性相关遗传 | |
| 第六节 人类的性染色体异常 |
第五章性别决定与伴性遗传
基本概念
初级性别决定是指性腺的形成。这可能受到特定基因的作用。性别的分化是指在性腺所产生的性激素的作用下,体细胞分化发育为内外生殖器和第二性征。
在单细胞真核生物中具有简单的性别决定系统,在此系统中由单对等位基因决定性别。在膜翅目昆虫中由一对杂合的染色体来决定雌性的发育。
不同的生物性别决定的机制不同,可分为四类,决定它们性别的主要因素分别是:性染色体、基因、性指数、环境条件如温度。但归根结底都和基因的调节有关。
三种不同的性染色体系统分别是XX-XO,XX-XY和ZZ-ZW。具有两个相同性染色体的性别称为同配性别,带有不同性染色体的成为异配性别。
大部分的植物都是两性的,少数具有XX-XY性染色体系统,成为雌雄异株植物。在女娄菜属中性别是由X和Y染色体的比来决定的。
和性别相关的遗传称为性相关遗传。性相关遗传有以下三种不同的类型:伴性遗传、从性遗传和限性遗传。
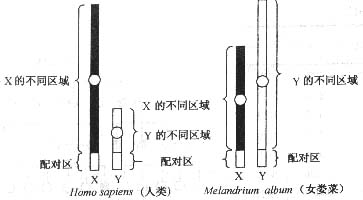
性染色体是由一对同源染色体进化而来的。在有的物种中两条染色体是同形的,有的物种中性染色体是异形的。前者属于原始的类型,后者属于进化的类型。在两条异形的性染色体之间只有很短的一段区域是同源的,称为假常染色体同源区。
性别也是生物的一种重要的性状,但比一般的性状要复杂得多。从远古到现在性别一直是人类探索的热点,它主要涉及到性别决定(sex
determination)和性别分化(sex differentiation)。前者指的是在各种因子的调控下,决定了性腺。后者指的是某些体细胞在性激素的作用下分化发育成内外生殖器以及第二性征。
一、 性染色体的发现
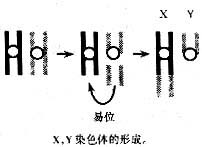
性染色体在高等真核生物的两种性别中是不同的。性染色体的发现为Sutton-Bovri的学说提供了一个实验证据。
在孟德尔定律第二次发现以前(1891年)德国的细胞生物学家亨金(Henking,H)曾经用半翅目的昆虫蝽做实验,发现减数分裂中雄体细胞着含有11对染色体和一条不配对的单条染色体,在第一次减数分裂时它移向一极,亨金无以为名,就称其为"X"染色体。后来在其他物种的雄性中也发现了X染色体。
二、 性染色体
和其他性状一样决定性别的不外乎是基因和环境两大因素,在有的生物中基因的作用是占主导地位,几乎不受外环境的影响;而另一些生物看来环境因素似乎起着主导作用。实际上环境仍然是通过基因在起作用。由于基因位于染色体上,因此在细胞学上所反映的常常是性别和性染色体之间的密切相关。和性别决定相关的染色体被称之为性染色体。
在一般情况下性染色体是由一对常染色体进化而来的,在进化时这一过程需要进行很多次。蛇就是一个很好的离子。人们研究了一些原始类型和高度进化的蛇的染色体。在原始类型的蛇中性染色体是相同的,称为同形(homomorphic)性染色体。这两条的差异仅仅在于复制的时期不同(在S期的晚期),实际上是W染色体。在较进化的种中,其中一条W染色体变小了,形成异形(heterochromatic)性染色体,这就是Z染色体。新莱昂(Lyon)假说认为:性染色体是同形性染色体部分易位到同源染色体上所至,所以Y染色体小于X染色体,而X染色体往往有重复的基因。低等脊椎动物的雄性和雌性个体的性染色体在形态上彼此是不能区分的。在它们的性染色体的某些区段上有许多等位基因的位点是两性共有的,而在另一些区段上两性所带的遗传信息却互不相同,雄性个体的性染色体所携带的遗传信息主要与性别决定有关。在鱼类中性染色体的分化就反映了这一进化趋势。
三、 性别决定
性别决定的本质是由一些相关基因的表达和调控来决定的,这方面的内容将在发育中予以讨论,现在还是从染色体和环境的角度来加以阐明。
我们可以把性别决定初步分为以下三类。
1. 性染色体决定性别
在这类中我们又可以分为6种不同的情况:
(1) 性染色体的本身来决定性别
XY型 在人类,哺乳动物,大部分的两栖类,爬行类,部分的鱼类和昆虫都属于XY型的。雄性为异配性别XY,雌性为同配性别XX。在这一类型中Y染色体起主导作用,不论X染色体有几条,只要存在1Y条染色体就发育为雄性。1990年辛克莱尔(Sinclair
,AH)等在前人的工作基础上发现在人和小鼠Y染色体的短臂上存在着性别决定基因,并在真兽亚纲动物中显示保守性。根据其在染色体上的位置命名为SRY(sex-determining
region Y chromosome),近年来又克隆出一系列与性别分化有关的基因,但SRY/sry是其中起主导作用的基因,因此携带此基因的Y染色体就成为决定雄性性别的标志。
在低等脊椎动物鸟类,两栖类,爬行类,鱼类以及某些植物中岁染也是由Y染色体来决定性别的,但机制尚不清楚。
ZW型
和XY型相反,在鸟类,鳞翅目昆虫以及大部分两栖爬行类等动物中是雌性异配性别(ZW),雄性是同配性别(ZZ),但性别决定的机制不像XY型那样研究得比较清楚。按照一般的推测,W染色体上可能也携带有和发育有关的基因或带有抑制雄性发育的基因。
(2) 性染色体的数目决定性别
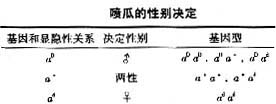
在某些双翅目,直翅目和鳞翅目的昆虫中没有异形的性染色体,而是由性染色体的数来决定性别。比如雌黄虫:2n=24(XX),雄性为2n=23(XO),比雌性少了一条染色体,称为XO型。植物中花椒也是属于XO型。在鳞翅目昆虫中也有雄性为ZZ,雌性为ZO的类型,称为ZO型。

1900年麦克朗(McClung,CE)等就发现了决定性别的染色体。他们采用的材料多为蚱蜢和其他直翅目昆虫。1902年麦克朗发现了一种特殊的染色体,称为副染色体(accessory
chromosome)。在受精时,它决定昆虫的性别。1906年威尔逊(Wilson,EB)观察到另一种半翅目昆虫(Proteror)的雌体有7对染色体,而雄性只有6条,另外加一条不配对的染色体,威尔逊称其为X染色体,其实雌性是一对性染色体,雄性为XO型。
在1905年斯蒂文斯(Stevens,N)发现拟步行虫属(Tenebrio molitor)中的一种甲虫雌雄个体的染色体数目是相同的,但在雄虫中只有一对是异源的,大小不同,其中有一条雌性也有,但成对的;另一条雌性中怎么也找不到,斯蒂文斯就称之为Y染色体。在黑腹果蝇中也发现了相同的情况,黑腹果蝇共有4对染色体,在雄性中有一条是异形的染色体(见图)。在1914年塞勒(Seiler)证明了在雄蛾中染色体都是同形的,而在雌蛾中有一对异形染色体。他们根据异形染色体的存在和性别的相关性,发现了性染色体,现在已经证实了他们的推论是完全正确的。
严格地说异形染色体的存在仅是一条线索,而不是证据,不能因为存在异形染色体,就表明其为性染色体。一定要通过实验证明这条染色体上存在决定性别的主要基因,方能定论。
新莱昂(Lyon)假说
3.
环境决定性别
在表型性别(phenotynic sex)决定系统中,有时环境起主要的作用。典型的例子是海生蠕虫后益(Bonellia),幼虫时没有性别的分化,自由独自在海底生活的幼虫发育成雌性,长约2英寸,有一条很长的吻部,其他的幼虫附着在雌体中寄居到雌体的子宫内,成为名副其实的寄生虫。这种蠕虫的性别不是受精时遗传成分所决定的而是由是否落在雌虫的吻部决定的。


(4) 染色体组的倍性决定性别
蜜蜂(Apis mellifera)的性别决定十分特殊,是由染色体组的倍性决定。蜂皇是可育的雌蜂,染色体为2n=32条,经正常减数分裂产生的卵为单倍体n=16,卵和精子(n=16)结合又形成2n=32的合子,将发育成蜂皇和工蜂(2n=32)。每一群蜜蜂中只有一个蜂皇,工蜂在遗传结构上和蜂皇并无差别,但由于工蜂所吃的蜂皇浆在质量和量上都远比蜂皇差,所以发育成不育的工蜂。蜂皇产生的部分单倍体的卵,少数未经过受精也能发育长成雄峰(n=16),雄峰的减数分裂十分特殊,第一次减数分裂时出现单极纺锤体,所以染色体全部向一极移动,两个子细胞中一个正常,含16个二联体,而另一个是无核细胞质芽体。正常的子细胞经第二次减数分裂产生两个单倍体(n=16)的精细胞,发育成精子。
(5) 取决于X染色体的是否杂合
小茧蜂(Habrobracon)的性别决定和蜜蜂又有不同之处,在自然状态下小茧蜂和蜜蜂相似,二倍体(2n=20)为雌蜂,单倍体(n=10)为雄峰。在实验室中,人们获得了二倍体(2n=20)的雄峰,其性别决定取决于性染色体X是纯合型还是杂合型。性染色体X有三种不同的类型:Xa、Xb、Xc。雌性的性染色体为一对杂合型的X染色体:XaXb或XaXc或XbXc;雄性为纯合型:XaXa、XbXb、XcXc。
(6) 取决于X和Y的比
大部分植物是没有性染色体的,多为雌雄同株,有的为雌雄同花,少数植物为XY型,称为雌雄异株植物。女娄菜的性别是由X染色体和Y染色体的比来决定的。酸模的性别决定又属于另一种类型,雌雄同株的植株为18A+XX+YY,X:Y+=1:1;雌雄异株中雌株为18A+XX+Y,X:Y=2:1;雄性植株为18A+X+YY,X:Y=1:2。但特殊的是,这两种交配型可以相互转换。具体机制尚不清楚。
2. 基因决定性别
(1) 由复等位基因决定性别
植物中有异形性染色体的并不多,但有的植物却明显由基因控制的,如葫芦科的一种喷瓜,是由复等位基因决定的。
(2) 由二对基因决定
玉米(Zea mays)一般都是雌雄同株的农作物。雌花的花序在叶腋呈穗状,由显性基因Ba控制的,其隐性等位基因ba。雄花的花序在顶端称为天花,由显性基因Ts控制,其基因型和表型性别的关系见表格。玉米的性别决定从图表中可见Ba只控制叶腋是否长雌花序。而Ts基因则不同,在它为显性时顶端长出雄花序,在其为隐性时不论顶端长出雌花序。也就是说不论另一对基因的状态如何,tsts存在时为雌雄,TsTs存在时顶端开雄花。
一、 外部的因素如温度、激素、感染
两栖类中如蛙是有性染色体的,XY型。蝌蚪在20℃时性别按其性染色体的构成长成分化。但在30℃时不论其基因型为XY还是XX,都发育成雄蛙。在爬行类中也是如此。
如扬子鳄和密西西比鳄的卵在不同的温度下,发育为不同的性别,当在30℃和30℃以下,发育为雌体;当温度在34℃或34℃以上发育为雄体,乌龟的卵在23-27的温度下发为雄性,在32-33时发育为雌性。所以他们卵产在不同高度的海岸线或河岸时由于温度不同而影响性别的分化,有可能是由于温度对两栖和爬行动物雌雄激素的合成有直接的影响。
在葫芦科的植物里,丰富的氮肥,短的日照和夜晚的低温也有利于雌性的发育。
二、 激素的影响
在哺乳动物中性别决定是指性腺的分化,性别的分化是指在性激素的作用下内外性器官的分化和第二性征的发育,毫无疑问性激素的作用是很大的。
在双胎牛中如果是一雄一雌,由于雄性的睾丸先分化,睾丸产生的雄激素通过绒毛膜血管流向雌性胎牛,使雌性胎牛的内外生殖器表现为雄性,但没有睾丸,呈间性。在人类中常见的一男一女的异卵双生子中为什么没有这种现象呢?那时由于人类异卵双生子是由两个胎盘将胎儿分隔开的,而牛的两个胎盘是共通的,由绒毛膜血管相连,所以可以相互影响。
三、 性反转的现象
生物从一种性别转变为另一种性别的现象称为性反转的现象(sex reversal)。这种情况有的是正常的自然反转,有的是人工引起的,或是环境条件改变所造成,或是由于病理因素所引起的。
酵母交配型的转变也属于性反转的现象,它是由基因控制的。在果蝇中存在一种性别转换基因(transformer)位于第3对染色体上。当tra雌果蝇的基因纯合时雌体发育成为不育的雄性。自然性反转在鱼类中有所发生,有一种海产鱼叫羊头鲷,幼小时鱼呈粉红色,为雄性,发育长大后变成黑色,性别转变为雌性。黄鳝(Monopterus
albus),属合鳃目,在我国仅一属一种。黄鳝染色体2n=24,皆为端着丝粒染色体,未发现异形性染色体的存在。黄鳝的体长和其发育阶段有一定的相关性,在个体发育过程中产生自然性反转。2龄前皆为雌性,3龄转变为雌雄间体,卵巢逐渐退化精巢逐渐形成。6龄全部反转为雄性。在欧洲鳗鲡,红鲈鱼及石斑鱼等的生活史也有性反转的现象。
有的自然性反转和上述相反,由雄性逐渐反转为雌性。如一种海产的蠕虫(Ophrgtrocha Puerilis)幼虫都是雄的,随着发育长大雄性生殖器退化,而雌性生殖器发育,比较"老"的个体都是雌的。但用人工的方法如饥饿,或在水中加入钾盐,或将其身体后部剪去可以促使雌体变成雄体。
牧鸡司晨的例子时有发生,有的母鸡停止产卵后竟然逐渐变成雄鸡,并能和雌鸡交配,但其基因型并未改变。
在人类中也存在着性反转现象,如46XX的男性和46XY的女性,这是由于SRY基因以及相关基因的异常引起的。
性反转不仅是表型的改变而且还涉及到性腺的改变。在生物中虽然有的有性染色体的分化,看来每一个体其性别是可以向雌雄两个方向分化的,也就是具备向两个方向发育的基因调节体系,外部条件或生理、病理条件的改变无非是影响了基因的调节而导致性反转。这个领域涉及到真核基因的调控,组织的凋亡,激素的分泌,组织的分化和发育,是十分复杂又非常有趣的。它已成为研究热点。
在20世纪初人们最初只发现交互的结果,产生相同的后代,并没有考虑到和性别之间有什么联系。第一个例外的情况是在1906年由唐卡斯特(Doncaster,L.)和雷纳(Raynor
G H)发现的,他们研究鹊蛾翅的颜色,用两种不同的品系:一种是亮白色(dw),一种是暗白色(DW),正交表明亮白为隐性;反交结果表明翅的表型和性别有关,雌性后代的表型和父亲相同,雄性后代的表型和母亲相同,如何解释这个结果呢?在解释前我们再考虑另一例子。





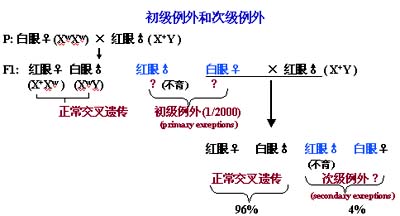
布里吉斯对唯一可育的初级例外(XXY)的减数分裂又提出了推理,分析有三种不同的分离类型,有84%是X-X染色体配对,然后相互分离,Y染色体随机地趋向一极;有16%的细胞是X-Y配对,然后相互分离,游离的Xw染色体也随机地移向两极,因此有一半(8%)将形成XwY和Xw型的子细胞。按这个假设初级例外形成的6种配子和正常的红眼雄蝇产生的两种精子杂交,将产生92%的正常后代和4%的次级例外。这是预料的情况,实验的结果获得了从以上比例看完全符合预料的结果,到此为止布里吉斯的模型比其他模型更具有说服力,他假设基因w或W在X染色体上,而且很好地解释了初级例外和次级例外,他的模型是受到了精确的检验的:
(1) 初级后代的细胞学研究表明雌性为XXY,雄性为XO,证实了布里吉斯的推论;
(2) 次级后代的细胞学研究表明雌性为XXY,雄性为XY,和推论相符;
(3) 例外白眼雌蝇的红眼女儿一半为XXY,一半为XX,和镜检结果一致;
(4) 例外白眼雌蝇的白眼儿子中也将产生例外的后代,这些白眼儿子都是XYY,此也同样得到了证实。
1916年摩尔根的三大弟子之一的布里吉斯(Brudges,G)发现了X染色体的不分离现象,才找到了染色体学说的直接证据。布里吉斯将白眼XwXw雌果蝇和红眼X+Y雄果蝇杂交时大部分后代是红眼雌蝇、白眼雄蝇,这与摩尔根实验的结果完全相同,但有1/2000的后代出现了意外的情况:红眼不育的雄蝇和白眼可育的雌蝇。他称其为初级例外后代(primary exceptional progeny)。他又进一步把初级例外的雌蝇和正常红眼雄蝇进行杂交,结果约为4%的后代是白眼雌蝇和可育的红眼雄蝇,他称其为次级例外,这是怎么发生的呢?


和性别有关的遗传都称为性相关遗传(sex-related inheritance),包括伴性遗传,限性遗传和从性遗传,三者特点是不同的。
一、 伴性遗传
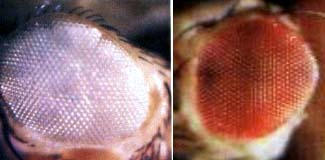
伴性遗传是指控制性状的基因在性染色体上,其遗传的特点由性染色体遗传规律所决定,基因的表达不受性激素的影响。
前面介绍的果蝇红眼和白眼的遗传就是伴性遗传,概括起来由个特点:正反交的结果不同;后代性状的分布和性别有关;常呈一种绞花式遗传(crisscross
heritance),即有害基因常常由母亲传给儿子,在儿子传给外孙女。
1. 人类的伴性遗传
人类中一些遗传性状或遗传病的基因位于X染色体上,由于Y染色体上只有少数的基因,一般没有X染色体上相应的等位基因,因此这些基因将随着X染色体一起传递。在女性中由于有两条X染色体,所以即使其中一条X染色体携带有致病的基因,由于是隐性的而得不到表达,我们称这种外在表型正常而携带有有害基因的个体为携带者(carrier)。在男性中只有一条X染色体,隐性的有害基因没有显性等位基因的存在,故称为半合子(hemizygote),这些隐性基因是可以表达的,正是由于这个原因显示出绞花式遗传。男性的平均寿命低于女性可能与此有关。据调查第一性比,即男性胎儿和女性胎儿之比为120:100,可能由于含有Y染色体的精子比含X染色体的精子更为活跃。第二性比是指男性婴儿与女性婴儿之比为(103-105):100,仍然是男性多于女性,但已低于第一性比;可能是在胚胎的发育过程中男婴的生命力比女婴弱;第三性比是指男青年与女青年之比为1:1,此表明人类的一夫一妻制的婚配制度正好是符合这一自然规律。也许是自然选择的结果。经统计85岁的老翁与老妪之比为62:100,相差悬殊,使人们不能不考虑到可能由于男性仅有一条X染色体的缘故。

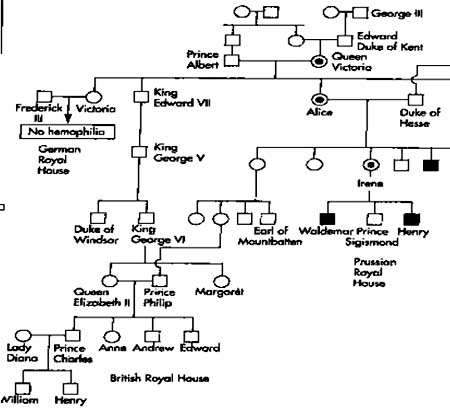
进行性肌营养不良(progressive muscular dystorphy,DMD),又叫假型肥大(pseudomu-pertrophic)或杜兴氏症(Duchemme's muscular dystorphy)表现为肌肉无力,走路缓慢呈鸭步,易摔到,上楼困难。仰卧起立时须先翻转为俯卧,再以双手支撑地面,然后再站起。常伴有腓肠肌、三角肌和冈下肌假性肥大。患者一般在12岁左右发病,20岁左右死亡。这个基因DMD已经分离出,为根治这种遗传病带来希望。
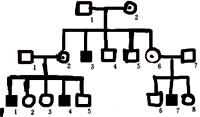
从典型的X-隐性遗传家系图谱可以看出以下特点:患者一般是男性;有害基因由母亲传递;呈绞花式遗传;双亲正常,儿子可以是患者;儿子若表型正常,后代都正常。

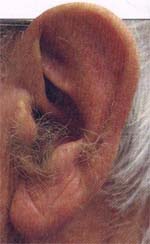
毛耳缘的表型和遗传谱系

典型的佝偻病的谱系
2. 植物的伴性遗传
植物中有雌雄同花,雌雄异花,雌雄异花中又有雌雄同株和雌雄异株之分,即使是雌雄异株也并不都有异形性染色体,如杨柳等。少数植物有异形性染色体分化,如女娄菜为雄性异配,草莓、金老梅为雌性异配。白剪秋萝也是雄性异配性别,叶片有宽叶和窄叶两种,将宽叶雌株和窄叶雄株杂交,F1无论雌雄全部都是宽叶,F1代互交得到的F2代雌性全部是宽叶,雄株半数是宽叶半数是窄叶,这个结果和果蝇的眼色遗传也完全相同。不难推论,控制叶形的基因在X染色体上,而且Y染色体上没有相应的等位基因。
| 物 种 | 常染色体 | 雄 | 雌 |
| 大麻(Cannabis sativa) | 20 | XX | XY |
| 茜草(Humulus lupulus) | 20 | XX | XY |
| 酸模(Runmex anglocarpus) | 14 | XX | XY |
| 女娄菜(Melandrium album) | 22 | XX | XY |

4. 不完全性连锁
在一对异形性染色体中,有部分区段是同源的,此段区域称为假常染色体同源区,减数分裂时可以联会。也有另一部分区段是不同源的,在减数分裂时不配对。位于X染色体不同源区域的基因和Y染色体不会发生交换,称为X-连锁基因,同样位于Y染色体上的非同源区域的基因就称为Y-连锁。位于X染色体和Y染色体同源区段的基因就称为X和Y连锁基因,由于这一区域有交换重组的可能,所以也称为不完全性连锁。如斯特恩(stern)发现果蝇的刚毛有一种突变型,刚毛变短,称截刚毛(bobbed),它的遗传如图所显示,其正反交的结果不是F1不同,而是F2不同,只能推论Y染色体上有相应的等位基因存在才可得到很好的解释。这也是唯一X和Y连锁的例子。


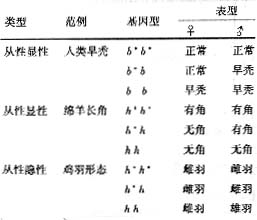
二、 从性遗传
从性遗传是指基因在常染色体上,但由于受到性激素的作用,基因在不同性别中表达不同,如人类头发的早秃(Baldness)这一性状就属于从性显性遗传(sex-influenced
dominance)。有的男子年纪很轻就开始秃顶,人们常误以为这是天资聪慧的标志,其实是一种遗传缺陷。带有这个基因(B)的杂合体只在男性中出现早秃的表型,在女性中表型正常,只有当B基因纯合时,女性才出现早秃,这个概率比带有单个B基因的概率小得多,所以很少女性是早秃的。绵羊是否长角的情况和以上相似。鸡羽毛是一个很好的从性隐性遗传的例子,雄鸡的羽毛和雌鸡不同,h基因控制雌鸡羽毛,只有当h基因纯合时在雄鸡中才能长出雄羽。从性遗传和X-连锁遗传不同,不是由X染色体传递的,而是由常染色体传递,另外表现出一定的基因型有时在雌性中不能表达,而在雄性都得到表达,可能是雄性激素的作用结果。其本质是基因的表达受到内部环境作用的结果。
三、 限性遗传
限性遗传(sexlimited inheritance)的基因可在常染色体或性染色体上,但仅在一种性别中表达,限性遗传的性状常和第二性征或激素有关。如毛耳的基因在Y染色体上,仅在男性中表达。睾丸女性化的基因位于X染色体上,但也只在男性中出现这种症状。子宫阴道积水也是一种遗传病,基因在常染色体上,但只在女性中出现。

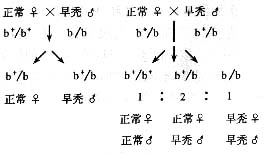
二、 克兰费尔特综合征(Klinefelter's
syndrome)
又称为"先天性睾丸发育不全"或"小睾症"。核型为47XXY,此是由于亲代减数分裂时,产生性染色体不分离所致,通过X染色体上Xg血型家系分析表现,60%的患者是由于母方,40%是由于父方染色体不分离所致。男性新生儿的发病率为1.3%,患者青春期后出现症状。身体瘦长,第二性征弱,常出现女性乳房,智力较差,睾丸小,精细管玻璃样变性,无精子发生,不育。
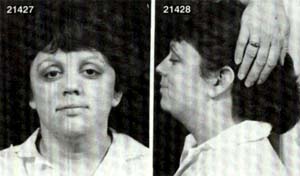
三、 特纳氏综合征(Turner's
syndrome)
又称为"先天性性腺发育不全"或"先天性卵巢发育不全"等。核型为45XO,X染色体阴性。1938年由美国H.Turner首先报道。在女孩中的发病率为0.2/1000-0.4/1000。患者表型为女性,第二性征发育不良,体矮(约为120-140cm)常有蹼颈和肘外翻,卵巢缺如,原发性闭经,无生育能力。智力低下,常伴有先天性的心脏病。产生的原因也是由于亲代配子形成过程中性染色体的不分离所致。约有75%的丢失发生在父方。约有10%的丢失了是在早期卵裂时。
Turner综合征患者发病远比Klinefelter综合征为低,这是由于约98%的45XO胚胎在早期已流产。只有约20%的个体发育异常较轻微才得以存活之故。
四、 XXY综合征
也称超雄,Sandberg(1961)首先报道,患者为男性,身材高大,少数有发育异常和智力较低,大部分核型为47XYY,X染色质阴性,有两个Y染色质,发病率占男性的1/750-1/1500。1965年Jacob等发现在暴力犯罪中XYY的比例较高(1:28),从而提出两个Y染色体的存在可能导致患者有侵犯行为。由于当时新闻界的渲染和扭曲,如把美国嗜杀成性的罪犯Speek..R描述成超雄。更引起了混乱。然而另一些研究者的报告指出XYY综合征患者的智力发育和社会行为完全正常,因此这种染色体异常对行为的影响尚无定论。
五、 XXX综合征
又称为超雌,Jacobs等在1959年首先报道。发病率为8/10000,多数患者表型和生育能力都正常。只有少数患者继发闭经,智力稍低等症状,细胞学检查可发现患者有2个Barr小体。患者母亲的平均年龄增高,表明染色体不分离现象主要发生在母方。
人类在减数分裂或在胚胎发育早期的有丝分裂中偶然会产生性染色体的不分开或丢失而造成合子的性染色体异常,这些个体也将会出现性别分化的异常。
一、 真两性畸形
真两性畸形不仅具有两性特征的内外生殖器,而且同时具有卵巢和睾丸两种不同的性腺,有40%的患者一侧有卵巢,一侧有睾丸;另40%的患者一侧为卵巢或睾丸,另一侧为卵睾(ovotestis);余下约20%的患者两侧均为卵巢。真两性畸形患者的副性征可为男性或女性,但其染色体却为嵌合型。主要分为两类:
(1) 46XX/47XXY;46XY/45XO;47XYY/45XO,这一类型是由于胚胎发育早期部分细胞有丝分裂过程产生了染色体不分离的现象,(如XYY/XO)或性染色体丢失(如46XXY/45XO)所造成的,这些细胞最初来源一个受精卵,所以称为同源嵌合体(mosaic)。
(2) 46XY/46XX,这种类型十分罕见,可能是由双受精的嵌合体发育而成。人类的嵌合体因起源不同分为两类:双胎嵌合体(twin
chimera)和双受精嵌合体(dispermic chimera)。到1975年为止全世界发现上述嵌合体各20例。Dunsford(1953)首先发现双胎嵌合体,其特点是有友伴双胎存在,双胎之间由于交互移植,出现混合血液;双胎一方对另一一方的移植的细胞缺乏凝集素,这是获得免疫耐受性的最好例子,这种嵌合体以自己的核型为主,他们具有正常的生育能力,我国在1983年也发现一例。Gartler(1962)发现另一类由双精受精引起的嵌合体。它是由两个精子和两卵核(可能为两个卵或一个双核卵,或一个卵,一个极体)受精后由于某种原因融合长成一个个体的结果,因而也称为整体嵌合体(whole
body chimerism),其特点是有混合型血液,体细胞培养物也显示镶嵌性。核型为46XY/46XX,表现为真性畸形,无生育力。
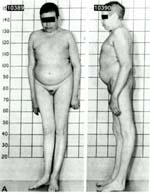


特纳氏综合征