


第四章 基因的作用及其与环境的关系
基本概念:
基因型要在一定的内外条件下才能表达为表型。环境对不同基因的表达影响不同。
一个基因如果存在多种等位基因的形式,这种现象就称为复等位基因(multipleallelism)。任何一个二倍体个体只存在复等位基因中的两个不同的等位基因。
在完全显性中,显性基因中纯合子和杂合子的表型相同。在不完全显性中杂合子的表型是显性和隐性两种纯合子的中间状态。这是由于杂合子中的一个基因无功能,另一个基因存在剂量效应。不完全显性中杂合体的表型是兼有显隐两种纯合子的表型。此是由于杂合子中一对等位基因都得到表达所致。
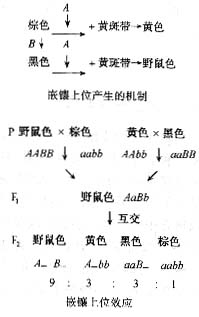
在很多情况下,由两对或多对基因通过对生化反应链不同步骤的控制,显示了非等位基因的互相作用。即一个基因的表型表达和另一对非等位基因有关,结果是孟德尔比发生变异。但对于二对基因来说,不论何种类型都是由(9:3:3:1)的变化而来。
非等位基因之间的"显性"作用称为"上位"效应,"隐性"作用称为"下位"效应。某些基因的等位基因表达时可导致个体死亡,这就是致死基因。若杂合致死则为显性致死,若纯合致死则为隐性致死。致死基因可能还控制别的性状,其显、隐性作用有时和在致死作用中的显隐性作用并不一致。
一个生物的基因型决定表型,但从基因型到一定的表型要在一定的环境条件下通过发育才能实现。那么环境或多或少对表型会有影响,同时表型是多种基因表型效应的共同作用的结果,基因间的相互作用又会对表型有什么影响呢?这一章将讨论环境作用对表型效应的影响以及基因间关系对表型效应的影响。
一、 遗传和环境
环境条件对多基因遗传的影响是十分明显的,如人的身高、胖瘦、肤色、智商等性状是受多基因控制的,营养状况、生活环境,受教育的情况对这性状都有直接的影响,即使是单基因遗传的质量性状也或多或少受到环境的影响。如半乳糖血症,是由于一个基因的突变使半乳糖苷酶缺失所致,但只要患儿不哺乳或不食用乳汁品就不会患病。水毛茛(Ranunculus
aguatilis)的叶片形态在不同的环境中是不同的。同一株植株,基因型完全一样,长在水下的叶片呈丝状,长在水面上的也呈掌状。藏报春(Primula
sincnsis)有的品种在20℃时花是红色的,而在30℃时花是白色的。喜马拉雅兔的毛色是受基因控制的在25℃左右时体温较低的部分,四肢和头部的尖端,尾巴和耳部的毛都是黑色的,其余部分全是白色。但在30℃以上的环境里长出的毛全是白色。如果把它躯干除去一部分毛再放到25℃以下的温度下,这部分新长出的毛是黑色的。当然生物有的遗传性状十分稳定,如人的血型,也是受基因控制的,基因型一旦形成表型就不会改变,不论生活在什么环境,营养状况如何,血型都不会改变。
|
第四章
基因的作用及其与环境的关系
|
| 第一节 环境的影响和基因表型效应 | |
| 第二节 显隐关系的扩展 | |
| 第三节 复等位基因 | |
| 第四节 基因间的相互作用与修饰 |
二、共显性
杂合子的一对等位基因各自都有自己的表型效应, 称为共显性(codominance)。MN血型是很好的例子。在人类的M-N血型系统中有三种血型,M,
N,MN型。他们是由基因型LM LM ,LNLN和LMLN决定的。M血型个体红细胞表面有M抗原, N型有N抗原,MN型有 M抗原+N抗原,当LM 、LN同时存在,都可以得到表现。也就是两种基因在同种组织中都得到了表达。这是由于一对等位基因中的一个发生了异效突变,它会产生不同的表型效应,当这一对等位基因杂合时,两种表型(M抗原和N抗原)同时存在。
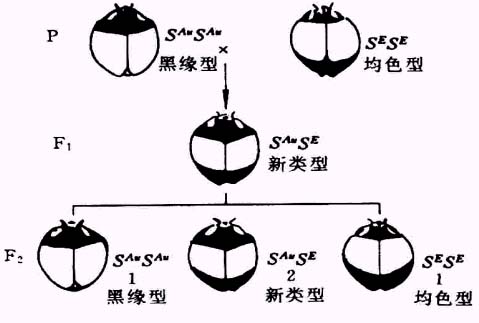
三、镶嵌显性
我国遗传学家谈家桢先生早年系统地研究了鞘翅瓢虫的遗传。发现了镶嵌显性。一种瓢虫鞘翅的底色为黄色,前缘呈黑色,称为黑缘型,另一种情况相反,鞘翅的后缘为黑色,称为均色型,两者杂交产生的F1代同时具有双亲的特点,即鞘翅前后缘都是黑色,这就是镶嵌显性。它和共显性是不同的,共显性是同一组织同一空间表现了双亲各自的特点,而镶嵌显性是在不同部位分别表现了双亲的表型镶嵌在一起。

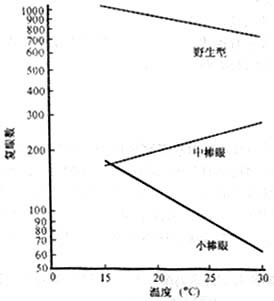


资料表明肺结核在不同人之间的共同发病率。肺结核是由结核杆菌引起的,但在群体中有人患病有人不患病,说明不同的人易感性不同,这种易感性是否由基因控制呢?
从资料中明显得出同卵双生子的同时发病率为87.3%,而异卵双生子的同时发病率为25.6%,差异是明显的;而异卵双生子和同胞之间的差别并不明显,说明肺结核易感性和遗传有关。
孟德尔在植物杂交实验中所观察到的7对性状都属于完全显性和隐性的关系。但并不是所有情况都如此。有时会遇到一些例子在显隐关系上出现各种变异。
一、不完全显性
不完全显性(incomplete dominance)又叫做半显性(semidominance),其特点是杂合子表现为双亲的中间性状。如紫茉莉,红花品系和白花品系杂交,F1代既不是红花也不是白花,而是粉红花;F1互交产生的F2代有三种表现型,红花,粉红花和白花,其比例为1:2:1。金鱼草的花色也是这样。安大路西亚(西班牙南部一个区域)鸡的羽毛,家蚕的体色,马的毛皮,金鱼身体的透明度等都属于此类不完全显性。



2. 不同环境条件对显隐性的影响
外部环境:
光和温度对生化反应有很重要的影响,这样也必然会影响到表型效应,以致改变显隐关系。如玄参科的金鱼草有红色和淡黄色两种不同的品系,如果将这两种不同品系杂交,所产生的F1代在不同的条件下表型不同。在光充足低温时F1为红色,那么红色是显性;当光不充足温暖条件下,F1为淡黄色,那么黄色为显性;当光充足温暖时呈不完全显性。可见外部环境条件会影响到显隐性的关系。
内部环境:
早秃属于内部环境影响显隐性的例子,Bb杂和的情况在男性中受到雄性激素的作用,单个的秃发基因可以表达,产生早秃性状,成为显性基因,而在女性中由于没有雄性激素所以不能表达,一定在有害基因纯合的情况下才能表达,从而表现为隐性性状。

在遗传学的发展早期就已经清楚,一个基因可能有两种以上的形式,然而一个二倍体生物在一个基因座位上只可能有其中的一对等位基因,而在一种生物群体中,某一个基因座位上的不同等位基因的总数常是相当大的。这种情况称为复等位基因(multiple
allelism),一套等位基因的本身就称为等位系列。等位性是遗传学中重要概念之一。所以我们要了解几个例子。



| 人类MHC四个位点的基因数和表型数 | ||||
| A | B | C | DR | |
| 复等位基因数 | 21 | 38 | 9 | 13 |
| 表型数 | 211 | 704 | 37 | 79 |


三、兔子的毛色
兔子的毛色也是由一组复等位基因构成的,C控制鼠色对其他3个复等位基因呈显性。Cch控制灰色,对Ch和c呈显性;Ch控制喜玛拉雅白化,对c为显性;c是一种突变型,不能形成色素,纯合时兔子毛皮为白色。
复等位基因所组合成的基因型数可以通过公式计算。

任何一个表型都是由多对基因控制的,现在我们来讨论两对非等位基因怎样来控制同一性状?他们相互间的关系是怎样的。根据孟德尔定律两对基因杂交其F2代的表型之比是9:3:3:1。前面讨论的是两个相互独立的性状,现在讨论的是同一个性状,显然情况不同,但万变不离其宗,仍然符合孟德尔法则,产生一些新的表型只不过是一种表型的综合而已。
在等位基因中有显隐之分,在两对非等位基因控制同一性状时也有"显"、"隐"之别,为了和等位基因的显隐相区分,人们就把两对非等位基因中起显性作用的称为上位基因(epistatic
genes),将起隐性作用的一对等位基因称为下位基因(hupostatic genes)或者说上位是指基因相互作用中一对基因的表型表达取决于另一对非等位基因的基因型。也就是异位显性。
一、 共上位
观赏鸟类有一种叫做虎皮鹦鹉的鸟,它的毛色有绿、蓝、黄、白四种,都是稳定遗传的,若将纯种的绿色和白色鸟杂交,或将蓝色与黄色杂交,F1代都是绿色的。F1代进行互交,F2代羽毛产生四种表型:绿、蓝、黄、白,比例是9:3:3:1。假设蓝色素是B基因控制的,黄色素是由Y基因控制的,B和Y都是完全显性的话,将黄鸟和蓝鸟杂交得到的BbYy是绿鸟,也就是黄色和蓝色的色素同时得到表达,混合在一起,我们看到的当然是绿色,如果B基因和Y基因都发生突变而不能形成色素,那么bbyy应当是白色,这样的假设正好符合虎皮鹦鹉的杂交。等位基因中有"共显性",那么在非等位基因中相似的现象我们称为共上位(coepistatic)。
六、 互补效应
香豌豆有紫色品系(PPCC)和白花品系(ppcc),杂交的F1代都是紫花(PpCc),自交后F2代只有紫花(P-C-)和白花(P-cc,ppC-,ppcc)两种表型,比例9:7,当然也是9:3:3:1的变形。也就是说只有当C、P都正常时才开紫花。这是由于紫色素的合成是经过了一系列的连锁反应,无论是C基因还是P基因发生突变,此反应都不能完成,而中间产物是白色的,所以只有C-P-基因型为紫色,其余基因型都是白色。这种类型称为互补效应(complement
effect)。和隐性上位不同的是其中间产物是没有产生色素(无色),在这种类型中C和P两个座位只要其中一个座位以隐性状态存在,对另一个基因都能起到异位显性的作用,故也称为双隐性上位(double
recessive epistatic)。





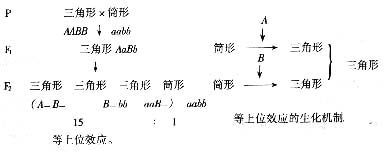
四、 等上位
荠的果形有三角形(AABB)和筒形(aabb)两种,三角形和筒形杂交F1代的果形都是三角形,F2代只有两种表型三角形和筒形,比例为15:1,这也是9:3:3:1的变形。从比例可知,除aabb以外,任何基因型只要哟一个A或B基因,表型都为三角形。A和B的作用相同,本身是完全显性,A和B互为完全显性,称为等上位(isoepistatic)或双显性上位(double
dominant epistatic),也就是剂量效应或重复基因(duplicate genes)。
图
五、 隐性上位
将黑色小鼠和白色小鼠杂交F1代是黑色,再将F1代互交产生的F2有三种表型,黑色,棕色,白色,比例为9:3:4。产生的原因是形成黑色素的过程是一个连锁反应。当C、B基因都正常时,才能产生黑色素,当B基因突变而C基因正常时虽不能产生黑色素,但能产生棕色的中间产物。当C突变即使B正常也因为缺乏中间产物而不能合成黑色素,反应停止在无色素原这一步,所以小鼠为白色。若C,B基因都突变,小鼠同样是白色。在这种类型中当C基因以隐性等位基因存在时,无论B基因是正常还是突变都不能发挥作用,似乎c对B、b起到异位显性作用,故称为隐性上位(recessive
epistatic)。


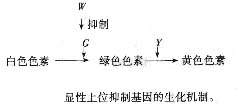
七、 显性上位
显性上位产生三种表型,比例是12:3:1,但产生这种比例的机制可能有不同的途径,一种是抑制基因引起的。在植物中有一种可以用于治疗心脏的药物叫毛地黄(Digitalis
purpuread)是一个很好的例子。毛地黄的M等位基因控制合成红色花色素苷。D为修饰基因,它控制合成大量的花色素苷,花呈深红色;d只能控制合成少量的花色素苷,花呈浅红。W基因座位控制花色素在花冠上的不同分布。W等位基因使红色色素只分布在筒状花的喉部,其余部位不分布。W基因使花色素分布于整个花冠。所以DdWw的表型为白色花喉部带红点,D-ww表现为既有大量的红色素又能分布于整个花冠,故呈深红色;ddww仅有少量的红色色素,也能广泛分布,花为浅红色。从以上可以看出,只有显性基因W的存在,无论是D还是d
,都是白色带红点,也就是说当W为显性时对另一个基因呈异位显性,也就称为显性上位(dominant epistatic)。
另一种类型的显性上位的分离比也是12:3:1。但机理不同,如南瓜的皮色有白色、黄色、绿色三种,遗传的
规律见图。产生的机制是由于W等位基因是一个抑制基因(inhibitor)作用的结果,原来产生黄色素要经过两步生化反应。当W基因存在时可以抑制G基因的表达,结果瓜呈白色。若W基因突变为w,则失去了抑制作用,反应可以顺利进行,直到产生黄色的色素,当Y突变为y时,则不能形成黄色色素,瓜呈绿色。结果也会同样出现12;3:1的分离比。
还有第三种可能的机制是深色和浅色同时表达,其结果是深色掩盖了浅色。如燕麦颖片颜色的遗传,当黑色颖片和白色颖片的品系杂交F1代为黑色颖片,F1代自交产生的F2代有三种表型,黑颖、黄颖和白颖,比例为12:3:1,产生的原因是黑色掩盖了浅色。B存在时,即使Y存在由于黑色产生掩盖了黄色
,表型仍然是黑色,只有b存在时Y表型为黄色,bbyy时既不能产生黑色素,也不能产生黄色色素,所以为白色。


|
两个位点供受体基因的排斥反应
|
|
移植组 --受体基因型-- 供体基因型 --反应 1 -------A1A2B5B5 ----A1A1B5B7 -排斥 2 -------A2A3B7B12 ---A1A2B7B7 -排斥 3 -------A1A2B7B5-----A1A2B7B7 -接受 4 -------A2A3B7B5 ----A3A3B5B5- 接受 |