现代学习遗传学的的学生们也许认为染色体学说并无惊人之处,但在20世纪初Sutton和Boveri的假说却十分轰动,人们对此学说发表的第一个反应是寻找它的漏洞,几年之后终于引发了一场激烈的争论。对Sutton-Boveri学说提出的某些异议是有价值的,而这些研究结果都证实染色体行为和基因行为是完全平行的。使得遗传的染色体学说更具魅力,但还没有直接的证据表明基因位于染色体上。直到发现性连锁才提供了直接的证据。






 精子的发生
精子的发生  卵子的发生
卵子的发生 遗传学的重要功绩就是解释了生物学中两大矛盾的生命现象“遗传”和“变异”。有丝分裂和减数分裂提供了重要的线索:有丝分裂是一个维持遗传结构的过程,而减数分裂却是产生变异的过程,通过独立分配和交换达到重组。
 中期Ⅰ
中期Ⅰ 早后期Ⅰ
早后期Ⅰ 晚后期Ⅰ
晚后期Ⅰ 末期Ⅰ
末期Ⅰ --间 期
--间 期 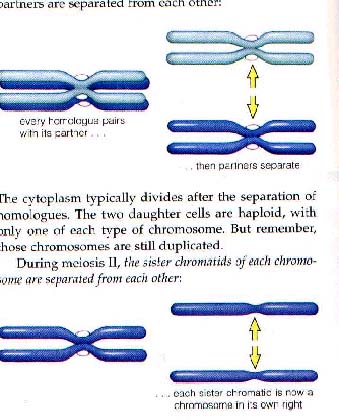
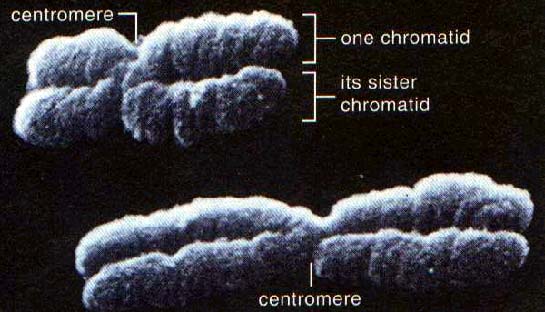
同源染色体分离和姐妹染色单体分离
 细线期
细线期
 偶线期
偶线期
 粗线期
粗线期 双线期
双线期
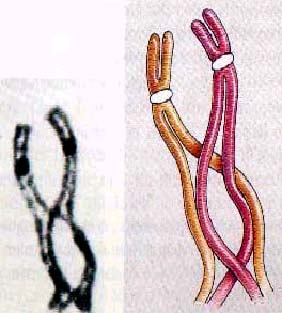 交 叉
交 叉  终变期
终变期
 前期Ⅱ
前期Ⅱ  中期Ⅱ
中期Ⅱ 后期 Ⅱ
后期 Ⅱ  末期Ⅱ
末期Ⅱ  四分体
四分体 花粉粒
花粉粒
动物细胞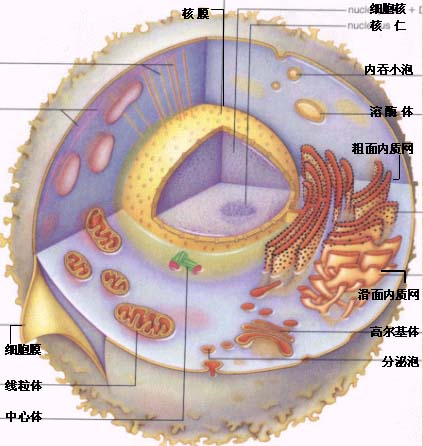
孟德尔分析的妙处就在于不知道基因是什么,或它们究竟怎样控制表型,而是用抽象的符号假设遗传因子来分析杂交结果和运用因子的分离和自由组合定律来预期杂交结果。在这个过程中不涉及任何关于基因的分子结构或是它们在细胞里的位置。然而人们还是会问:究竟细胞中什么结构与假设的遗传因子(基因)一致?
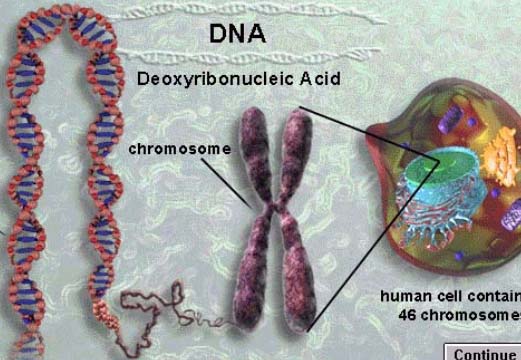
遗传学发展的重要里程碑是由于接受了这样一个概念,就是:被孟德尔研究过的决定遗传特性的基因是与作为细胞的特殊结构的一部分--染色体相关联的概念。细胞学提供了在显微镜下可以看到的染色体的行为与杂交实验的遗传因子的行为相关联。这一重要概念来自遗传的染色体学说。它标志着遗传学与细胞学的结合。直到今天,这一学说仍然是遗传分析的本质的一部分,在医学遗传、农业遗传和进化遗传上有重要的应用意义。
认识遗传的染色体学说,必须掌握细胞分裂过程中的染色体行为及其相关的概念。
第 一 节 细 胞的结构
细胞是多细胞生物最小的“建筑”单位,细胞只可能由另一个细胞通过分裂产生。
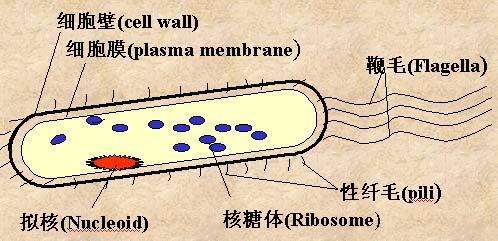
原核生物是最简单的单细胞生物,如细菌。原核生物缺乏由膜包被的核,以二等分裂(binary fission)来繁殖。原核生物的遗传物质是单个的环状DNA分子,DNA分子上结合有少量的蛋白质。真核生物的细胞中,遗传物质位于有膜包被的核中,遗传物质分布在各条线状染色体上,每条染色体都由很多蛋白质和DNA组成。
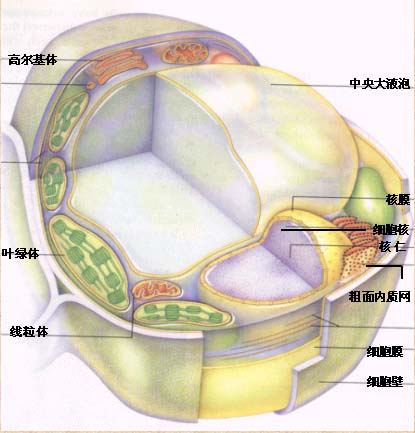
关于细胞的各种结构与功能的具体情况请参阅细胞生物学。
真核细胞结构:
 间
期
间
期  早前期
早前期  晚前期
晚前期 中 期
中 期  后 期
后 期  末期
末期
19世纪末Flemming W(1882)和Boveri(1891)分别发现了有丝分裂(mitosis)和减数分裂(meiosis),为遗传的染色体学说提供了理论基础。在孟德尔研究和孟德尔定律的再发现之间(1886-1900)有很多生物学家并不知道孟德尔的发现,但对遗传现象很感兴趣,而且用完全不同的方法来研究这个问题。这些研究者想知道遗传物质在细胞的什么部位。很显然值得寻找的地方是配子,因为它们是亲子之间唯一的联系。卵和精子的大小不同,卵细胞质的量很多,而精子很小,其细胞质不像是遗传物质的所在地,卵和精子的核大小相同,因此被认为是遗传结构比较合理的藏身之地。细胞核的主要成分是什么?很明显是染色体,它在所有的细胞中都存在。这一特点强烈吸引着生物学家,他们关注的是在一个生物体中不同细胞之间染色体数目的恒定性。那么染色体数目的恒定性是怎么维持的呢?要回答这个问题只有在有丝分裂和减数分裂时用显微镜观察染色体的行为。从观察的结果中发成遗传的染色体学说--染色体上带有基因。
一、有丝分裂
有丝分裂:是二倍体或单倍体真核细胞的分裂方式,其特点是在分裂的过程中出现染色体丝和纺锤丝,分裂结果是形成两个具有相同遗传物质的新细胞。
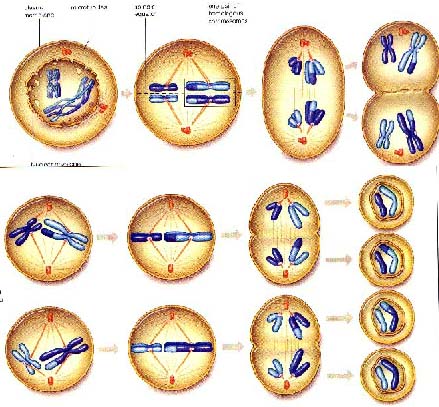
细胞周期:(cellcycle)是从细胞的任何一个分期阶段到子细胞的同等阶段的各种事件。或从一个新产生的细胞到它分裂产生子细胞的这一过程称为细胞周期,它可分为四个阶段:M期、S期、G1期和G2期。M期是细胞周期中最短的时期,约占整个时期的5-10%的时间。DNA的合成在S期(synthesis)。G1(gap1)期和G2(gap2)期是S期和M期之间的两个间隙期。G1、S、G2合称为间期,即两个分裂期之间的时期。此期染色质均匀分布于核中,所以在显微镜下看不到染色体。染色质网形成时核中的染色体完成了复制,然后再纵裂分开形成两条染色体,每一条进入不同的子核中,有丝分裂产生了两个完全相同的子核,分裂成两个子细胞。为了便于研究科学家们把这一连续的过程划分为前期(prophase)、中期(metaphase)、后期(anaphase)和末期(telophase)四个阶段。
有丝分裂过程图解(百合根尖细胞有丝分裂图象)
前期:染色体开始逐渐变得清晰可辨,逐渐凝集缩短变粗,收缩成螺旋状,这种形状易于移动。每条染色体已含有两条染色单体(chromatids),互称为姐妹染色单体(sister chromatids)。通过着丝粒(centromere)相互连接在一起。前期末核仁(nucleoli)逐渐消失,核膜开始破裂,核质和细胞质融为一体。
中期:纺锤体(spindle)逐渐明显,这个鸟笼状的结构在核区形成,有细胞两极间一束平行的纤丝构成。着丝粒附着在纺锤丝上,染色体向细胞的赤道板(equatorial plane)移动。
后期:着丝粒纵裂为二,姐妹染色单体彼此分离,各自移向一极。染色体的两臂由着丝粒拖曳移动,这时染色体是单条的,称为子染色体。
末期:末期子细胞的染色体凝缩为一个新核,在核的周围核膜重新形成。染色体又变为均匀的染色质,核仁重新出现,重新又形成了间期核。末期结束时纺锤体被降解,细胞质被新的细胞膜分隔成两部分,结果产生了两个子细胞
二、减数分裂
减数分裂发生在有性生殖的真核细胞中。它是由二倍体细胞或核通过一次DNA复制和二次分裂产生4个单倍体细胞或核的分裂过程。
减数分裂特点:
1.具有一定的时空性,它仅在一定的发育阶段在生殖细胞中进行。而有丝分裂几乎在生物的一生中在各种不同的组织中都进行。
2.减数分裂经第一次分裂后染色体就进行减半,其产物是单倍性的。
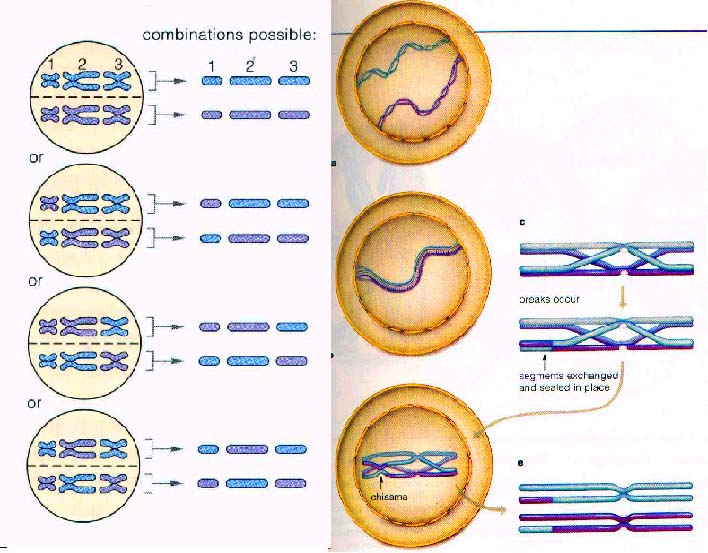
3.前期长而复杂,同源染色体经历了配对、联会、交换,从而使遗传物质进行了重组。
4.每个子细胞遗传信息的组合不同。
减数分裂过程图解(花粉的形成)
减数分裂Ⅰ:前期Ⅰ(prophaseⅠ)
细线期(leptotene):此期染色体呈细线状,核仁依然存在。在细线期和整个的前期中染色体持续地浓缩。细线期中沿着每条染色体浓缩的小区域称为染色粒(chromomeres),呈链珠状。
偶线期(zygotene):细线状的染色体开始配对,在性母细胞中,实际上有两套染色体,每条染色体都有一条与之同源的染色体,逐步相互配对或联会(synapsed),就像“拉链”式的两个成员称为同源染色体(homologs)。两条同源染色体精确配对的机智是怎样的呢?虽然这个机制目前还没有完全弄清楚,但我们已经知道需要一种蛋白质和DNA构成的精巧结构,称为联会复合体(synaptonemal
complex,SC),联会时它总是夹在两条同源染色体之间。
粗线期(pachytene):在这一阶段,染色体完全联会,缩短变粗,但核仁依然存在。在核中同源染色体的对数等于n。一对配对的同源染色体称为二价体或四联体,在特殊情况下,存在不能配对的染色体则称为单价体或二联体。染色粒的直线排列在每一对的同源染色体上,像一串精致的珠链。
双线期(diplotene):在细线期时每条同源染色体看起来都是单条的线状,其实此时DNA已在S期复制过了,只不过难以分辨出而已。显然到了双线期每条染色体就出现两条,与有丝分裂中期相似,其中每一条就称为染色单体。由于配对的同源染色体每条都产生两条姊妹染色单体,所以联会复合体的结构是一束四条染色单体,在双线期,同源配对时不太紧,其实是相互排斥呈现轻微的分离。非姊妹染色单体间的交错结构称为交叉,每一对同源染色体都有一个或多个的交叉存在。发生在减数分裂早期的交叉称为交换(crossover),可以发生在偶线期或粗线期。交换主要发生在减数分裂,在有丝分裂中十分罕见。一个交换是两个非姐妹染色单体之间的一次精确的断裂、互换和重接。每对染色体至少发生一次交换,这对于正常的分离是必要的。交换的另一个作用是产生新的基因组合,这是群体遗传变异的一个重要来源。交叉的端化,非姐妹染色单体之间要彼此滑动,这样就延缓了染色单体的分离。
浓缩期或终变期(diakinesis):此期明显不同于双线期,染色体内进一步地收缩,常可见到“O”形或“+”的一对同源染色体,这是端化的结果,染色体的收缩便于分裂时移动。
中期Ⅰ:核膜、核仁消失,每对同源染色体位于赤道板上,着丝粒分居于赤道板的两侧,附着在纺锤丝上。而有丝分裂的中期着丝粒位于赤道板上,中期Ⅰ着丝粒不分裂。这一点和有丝分裂明显不同。
后期Ⅰ:在有丝分裂中当染色体向两极移动时后期开始了。在减数分裂的后期Ⅰ是一对同源染色体彼此分离,向相对的两极移动。
末期Ⅰ:此末期和随后的“间期”也称“分裂间期”(interkinesis),并不是普遍存在的,在很多生物中没有这一阶段,也没有核摸重新形成的过程,细胞直接进入第二次减数分裂。在另一些生物中末期Ⅰ和分裂间期是短暂的,但核膜重新形成。在很多情况下,此期不合成DNA,染色体的形态也不发生改变。
按照惯例,染色体到达两极,解旋后成为染色质,重新形成两个子核,每一个核是一个单倍体,这是因为它只含有一套染色体,但每一条染色体都含有两条染色单体附着在着丝粒上。也就是说我们可以以着丝粒点来计算染色体条数,而不管是否有姐妹染色单体的存在。在每一个子细胞中染色体减半是个关键,因此减数分裂Ⅰ被称为“减数分裂”,而第二次减数分裂和有丝分裂相同,染色体的数目保持不变,这种类型的分裂称为“等数分裂”。
减数分裂Ⅱ
前期Ⅱ:此期和前期Ⅰ的情况相似,每条染色体都已经复制过,所不同的是此期染色体数是n。
中期Ⅱ:染色体排列在赤道板上,纺锤丝附着在单个的着丝粒上。染色单体从彼此紧密相连逐渐部分地分离。
后期Ⅱ:着丝粒纵裂,姐妹染色单体由纺锤丝牵引向两极移动。
末期Ⅱ:在子细胞两极染色体周围核膜重新形成。
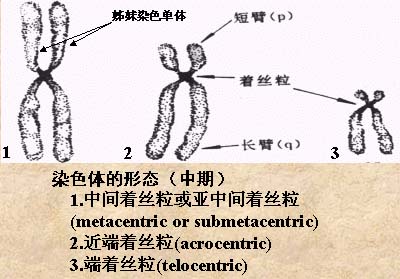
三、染色体:
在细胞分裂过程中出现的一种具有一定形态、数量、能被碱性染料着色的携带有遗传信息的小体。形态多种多样,如棒状、颗粒状;没有生物膜。
一、动物的生活史
绝大多数多细胞动物在其大部分生活史中都是以二倍体存在的。在每种动物中,经过减数分裂产生单倍体配子。通过受精不同配子的核进行融合再形成二倍体的合子。经过不断地有丝分裂发育成一个新的二倍体生物。这样配子仅是生命周期中的单倍体阶段,配子仅在特殊的组织中发现。雄性动物产生的配子是精子。这个过程称为精子发生。雌性的配子是卵,由卵子发生而形成。
在雄性动物中精细胞是由睾丸产生的。睾丸中含有原生殖细胞,也叫初级精原细胞,通过有丝分裂产生次级精原细胞,由它转变成初级精母细胞,每个精母细胞经过第一次减数分裂形成两个次级精母细胞,经过第二次减数分裂每个次级精母细胞形成两个精细胞,这样一共产生4个单倍体精细胞。经过进一步分化成成熟的雄配子,即精子。
在雌性动物中卵巢含有初级卵原细胞,通过有丝分裂产生次级卵原细胞,这些细胞再转化成初级卵母细胞。初级卵母细胞通过第一次减数分裂和不均等的胞质分裂产生两个细胞,大的一个细胞称为次级卵母细胞,很小的一个称为第一极体。经过第二次减数分裂,次级卵母细胞产生两个单倍体细胞,一个是很小的细胞称为第二极体,另一个较大的细胞迅速地发育为成熟的卵细胞。第一极体可分裂可不分裂。只有卵子是有生命力的配子。这样在雌性动物只能产生一个成熟的单倍体的卵。
二、植物生活史
在具有有性生殖生活周史的植物中有两种典型的时期。一种是配子体时期或单倍体阶段,在此阶段产生配子;另一种是孢子体时期或二倍体阶段,此期产生二倍体孢子母细胞。
在被子植物中显花植物的花是有性生殖器官,通常含有雌雄两种生殖器:雄蕊和雌蕊。雄蕊含有单个的柄,花丝在柄的顶端也就是花药。花药里有很多花粉,这是重要的雄配子体。雌蕊含有雌配子体及典型的柱头,表面粘着,易于接受花粉。
减数分裂的一种特殊的分裂方式在花的雄蕊和雌蕊中进行。在花的子房中,每一个胚珠是一个大的2n细胞,称为大孢子母细胞,这些细胞经过减数分裂产生两个单倍体产物,其中一个保持了生命力,这就是大胞子,或单倍体雌配子体。大孢子的产生叫做大孢子发生。
减数分裂在大孢子母细胞中通常涉及两次分裂,产生4个单倍体细胞。其中3个退化,只有一个大细胞能生存下来,它具有一个单倍体核,再经过3次有丝分裂,大孢子体带有8个相同的单倍体核,这8个单倍体核在细胞的两端,每端有4个。然后每端有一个核回到细胞的中间部位。接着细胞壁产生,导致细胞质的不均等分裂,中间的两个核的周围形成了大的双核细胞,受精时和一个雄核结合产生3倍体胚乳,因此称它为胚乳母细胞或极核。其他6个核各自被封闭在细胞特定位置上。整个具有8核的位于靠近胚乳的端部的大配子体称为胚囊。6个单倍体细胞中的一个称为卵细胞,它将成为胚珠中单个的雌配子体,它随机和一个精细胞结合而受精。
产生花粉粒或雄配子体也要经过减数分裂,但要简单一些,这个过程称为小孢子的发生。花药含有4个花粉囊,其中有很多小孢子母细胞。每个小孢子母细胞经减数分裂产生4个单倍体小孢子。每个小孢子经过一次有丝分裂产生一个单倍体的生殖核和一个单倍体的管核,然后产生一个硬壳成为花粉粒。
受精时花粉粒附着在柱头上萌发后产生花粉管。花粉管生长穿过柱头进入花柱,在花粉管生长前产生的酶使组织降解,当花粉管伸入花柱,核停留在生长的顶端;单倍体生殖核再经过一次有丝分裂产生2个精核或雄核。此时管核不再分裂。花粉管最后穿入胚珠上的一个小孔叫珠孔,两个精核进入胚囊,一个精核和单倍体卵细胞受精,产生二倍体合子;另一个精核与两个极核融合,产生一个三倍体核,这就叫双受精(double
fertilization)。双受精仅在显花植物中存在。
关于遗传的染色体学说,这个概念是把基因看作染色体的一部分。这是由一位美国年轻的毕业生Walter Sutton和一位德国的生物学家T.Boveri提出的。1902年孟德尔定律重新发现后不久,引起人们的极大兴趣,他们独立地认识到豌豆产生配子时孟德尔因子的行为和减数分裂时染色体行为有着精确的平行关系。在真核生物中基因是成对存在的。人们称之为等位基因,而染色体也是成对存在的,称为同源染色体;在形成配子时等位基因分离分别进入不同的配子中,一对同源染色体在减数分裂第一次分裂时也分开,移向细胞两极;不同的基因在形成配子时自由组合地进入配子,非同源染色体也是如此。这两位研究者通过分析得到相同的结论。