 |
第十三章遗传工程
|
| 第一节 基因工程的原理和一般方法 | |
| 第二节 限制性内切核酸酶与连接酶 | |
| 第三节 基因工程中的载体 | |
| 第四节 基因组克隆和cDNA文库 | |
| 第五节 转基因技术及其应用 |
基因工程(gene engineering)是20世纪70年代初期诞生的一门崭新的生物技术科学。它是指采用类似工程设计的方法,人为地在体外将核酸分子插入质粒、病毒或其他载体中,构成遗传物质的新组合,并将它转移到原先没有这类分子的寄主细胞中扩增和表达。
|
第十三章遗传工程
|
| 第一节 基因工程的原理和一般方法 | |
| 第二节 限制性内切核酸酶与连接酶 | |
| 第三节 基因工程中的载体 | |
| 第四节 基因组克隆和cDNA文库 | |
| 第五节 转基因技术及其应用 |
基因工程(gene engineering)是20世纪70年代初期诞生的一门崭新的生物技术科学。它是指采用类似工程设计的方法,人为地在体外将核酸分子插入质粒、病毒或其他载体中,构成遗传物质的新组合,并将它转移到原先没有这类分子的寄主细胞中扩增和表达。
第一节 基因工程的原理和一般方法
一.基因工程的基本原理
从基因工程的定义出发,它的第一个重要特征就是可把来自任何一种生物的基因放置到与其他毫无亲缘关系的寄主生物中,因而应用基因工程技术,就可能按照人们的主观愿望,改造生物的遗传特性乃至创造出自然界中原本不存在的新的生物类型。第二个特征是它表明一种已知的DNA片段在新的寄主细胞内可以进行扩增。这样就为制备大量纯化的DNA片段提供了可能,从而拓宽了分子生物学的研究领域。
二、基因工程的基本内容和技术
(一)基因工程的基本内容
基因工程是把来自不同生物的外源DNA插入到载体分子上,以形成“杂种”DNA分子,然后将之引入寄主细胞进而实现目的基因功能的表达。基因工程的核心技术是分子克隆(molecular
cloning),其次是外源基因的高效表达,而基因表达也离不开分子克隆技术。概括起来,基因工程的主要内容大致是:
1.从复杂的生物体基因组中,经过酶切消化等步骤,分离带有目的基因的DNA片段,或用酶学和化学方法人工合成基因。
2.将外源DNA片段与能够自我复制并具有选择标记的载体分子在体外连接,形成重组DNA分子。
3.把重组的DNA分子引入到适宜的受体(寄主)细胞中进行扩增。
4.从繁殖的大量细胞群体中筛选和鉴定,以获得重组DNA分子的受体细胞的克隆。
5.从所筛选的受体细胞克隆提取已扩增的目的基因后,或再将其克隆到表达载体上,导入寄主细胞,以便在新的背景下实现功能表达,产生人们所需的物质;或是将已扩增的目的基因作进一步的分析研究。
(二)基因工程的基本技术
20世纪70年代初期基因工程的出现并不是一种偶然的事件,而是由科学技术的发展水平所决定的。如在核酸研究方面,采用酶学、新的柱层析和超速离心的技术等改进了核酸的提取方法,能保证获得分子大而完整的纯净样品;并建立了如电泳、酶切图谱分析、分子杂交、电子显微镜技术等新的微量鉴定方法,以及DNA序列测定和基因的人工合成等多种分子生物学实验方法,从而为基因工程的创立奠定了强有力的技术基础。
基因工程操作的主要步骤是:分离或合成目的基因,将带有目的基因的DNA片段与载体DNA体外重组,然后把重组体转人受体细胞以及重组体克隆的筛选和鉴定。但在具体的基因工程操作中则还应根据实验目的、基因片段的来源与性质来选择相应的技术路线。
一.限制性内切核酸酶的类型
限制性内切核酸酶(restriction endonuclease)常简称为限制酶(restriction
enzyme),是一类能识别双链DNA分子中特异核苷酸序列并由此切割DNA双链结构的水解酶。
这些限制酶有着共同的特性,如它们只降解双链DNA分子,不切单链DNA;各种酶有其特定的核苷酸序列识别特异性;酶的活性需要Mg2+离子来激活等等。但不同的酶也有着许多差别,根据它们自身的结构。切割位点和识别序列间的距离不同。使酶激活所需要的辅助因子的差异以及与甲基化作用的关系等,可将限制酶分为I、Ⅱ和
Ⅲ型。
一.分子克隆的载体及其所应具备的基本条件
目前所用的载体有细菌的质粒(如大肠杆菌质粒)、噬菌体或病毒,更多的是经过人工改造的一些载体。用于分子克隆的载体都应具备下述条件:
① 在细胞内能自主复制,即具有复制原点,可以使所携带的外源DNA在受体细胞内忠实地复制;
② 有适宜的限制酶切位点。最好是对多种限制酶有单一切点,且酶切位点不在复制原点内。
③具备对重组体易于检测的选择性遗传标记。
除上述的基本条件外,根据实验的需要,人们还常希望载体具有其他的各种特征。如携带外源DNA的幅度较宽,具备可促进外源DNA高水平表达的调控区;为保证安全,载体在特定宿主细胞外不能生存,以及某些特殊的筛选指标等等。
根据DNA分子克隆的目的,载体可分为:
克隆载体:是指以繁殖DNA片段为目的一类载体。这类载体一般较小,其复制不是处于寄主的严谨控制下,因此特定的DNA序列可以通过克隆技术大量制造,如pBR322
穿梭载体:是指那些既能在真核细胞中繁殖又能在原核细胞中繁殖的载体。这类载体必须既有细菌的复制原点或质粒的复制原点,又含有真核生物的复制原点,还具备酶切位点和合适的筛选指标。它用来转化细菌,又可以用于转化真核细胞。
表达载体:是用于使克隆的外源基因在寄主细胞内表达的一类载体。它要有很强的能为寄主RNA聚合酶识别的启动子,有使RNA聚合酶只转录克隆的基因而不能转录无关序列的强终止子;其次是启动子还应是受控制的即只在诱导时才进行转录,以保证转录效率,并防止给细胞造成不利影响。
二.常用的载体
(一) 质粒载体
质粒是细菌细胞中独立于细菌“染色体”之外能自主复制的遗传单位。质粒DNA分子在宿主细胞内可以持续稳定地游离于染色体外,但在一定条件下又能可逆地整合到宿主染色体上并随之复制,通过细胞分裂传递到后代。质粒DNA是共价闭合的环状双链分子,常带有一些特殊的基因,如致育基因,合成抗菌素的基因、抵抗抗生素的抗性基因以及降解复杂有机化合物的基因等,从而赋予寄主细菌一些额外的特性。
质粒具有不相容性(incompatibility),就是指在没有选择压力的情况下,两种亲缘关系密切的不同质粒不能共存于同一宿主细胞内。在细胞增殖的过程中必定有一种会被逐渐排斥掉。这样的两种质粒称为不亲和质粒。正是基于质粒的不相容性,分子克隆可在只带一种质粒的细菌中提取单一的质粒DNA供作载体进行
DNA重组。
(二)噬菌体载体
1. λ噬菌体载体
λ噬菌体是迄今为止研究得最为详尽的一种大肠杆菌双链DNA噬菌体。它的DNA分子两端各有由12个核苷酸组成的粘性末端,两者互补可形成COS位点(cohesive
end site)。
2. 柯斯质粒载体
柯斯质粒(cosmid)是人工构建的带有粘性末端cos的一类质粒,其主要组成有质粒的复制起始区和抗药标记、一种或多种限制酶的单一切点以及λDNA的COS区小片段(约20至数百个碱基对)。因而这类质粒兼具λ噬菌体和质粒的优越性,又具有高容量的克隆能力,容纳外源DNA的长度可达45
kb左右。
应用柯斯质粒为载体,在大肠杆菌细胞中克隆基因组大片段DNA的技术称为柯斯克隆(cosmid cloning)。这一技术的优点是,首先由于柯斯载体的前述特性,提高了克隆外源DNA片段的能力,是构建真核生物基因组文库特别有用的克隆载体;其次,以柯斯质粒为载体,所形成的非重组体的克隆其本底常较低。
(三)人工染色体
以噬菌体为基础构建的载体以及cosmid(粘粒)系统只能接纳在一定大小范围内的插人片段,只是一些真核基因过于庞大不能作为单一片段克隆到这些载体中。于是发展出来
P1噬菌体人工染色体(p1artificial chromosome,PAC)、细菌人工染色体(bacterial artificial chromosome,BAC)、酵母人工染色体(yeast
artificial chromosome,YAC)等新的克隆技术。其中酵母人工染色体克隆系统已较完善。
以酵母作为构建人工染色体的材料,是由于其基因组很小,约为1~14mb,且其中极少内含子,没有重复序列和假基因。其基因行为和表达方式基本上与高等生物类似,如复制、重组、染色体分离等等,因而是研究真核生物分子遗传学的重要实验模型。加之近年对有关酵母染色体的主要组分:着丝粒、自主复制序列(autonomously
replicating sequence,ARS)及端粒的研究又较清楚,并被分离和克隆出来,其功能区已缩小到数百碱基对,这些对研究高等生物染色体的结构与功能也是极为有利的。
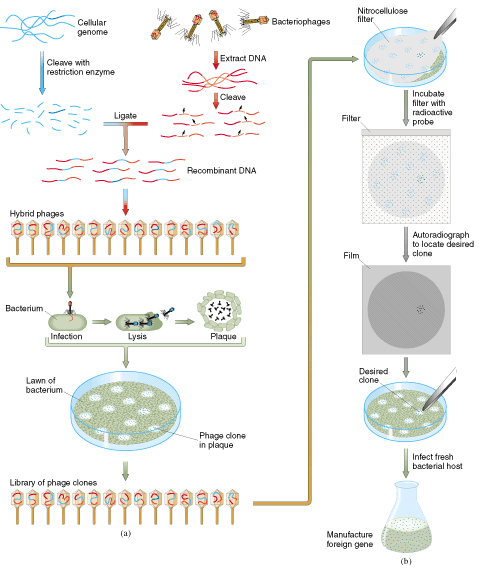
一.基因组克隆
用限制性内切酶切割整个基因组DNA,把所得的大量基因组DNA片段与载体连接,然后转化到细菌中去,让宿主菌长成克隆。由此而得的克隆中每个细胞的载体上都包含有特定的基因组DNA片段。这样的一套克隆便称作基因组克隆,而这些克隆中的一套基因组DNA片段则叫做基因组文库(genomic library)。
二.cNDA 文库的构建及应用
cDNA基因文库(cDNA library)是指以 mRNA为模板,在反转录酶及其他一系列酶的催化作用下所获得的双链cDNA,经与适当载体连接并转化到寄主细胞内进行扩增,由此而构成包含着相应基因编码序列的一群克隆。与基因组DNA克隆不同,用作cDNA克隆的真核mRNA,其间隔序列早已在拼接过程中被删除,而由这种成熟mRNA为模板转变而来的DNA基因,自然也不含有非编码序列。cDNA克隆除以
mRNA为起始材料这一优点外,其构建比较简易可行,而且由于每一个cDNA克隆都只含有一种mRNA序列,这样在选择中出现假阳性的概率也就较低。
随着分子克隆研究在理论和技术上的进步,cDNA 基因文库的构建不仅是分离基因的重要手段,也是当前真核分子生物学研究的强有力工具。首先,在基因工程的操作中,以cDNA为探针可从基因组文库中分离出相应的特异基因;其次,cDNA序列只与基因中的编码序列有关,有助于对真核生物mRNA结构和功能的认识;再者,cDNA克隆还可以有效地分析真核基因的结构和功能。
特 性 |
I 型酶 |
II 型酶 |
III 型酶 |
限制和修饰活性 |
单一多功能的酶 |
单一的成分 |
2种不同的亚基 |
蛋白质结构 |
3种不同的亚基 |
单一的成分 |
2种不同的亚基 |
辅助因子 |
ATP、Mg2+、S-腺苷甲 |
Mg2+ |
ATP、Mg2+、(S-腺 |
寄主特异性位点序列 |
EcoB:TGANTGCT |
旋转对称 |
EcoP1:AGACC |
切割位点 |
距特异性位点至少 |
位于特异性位点 |
距特异性位点 |
DNA易位作用 |
能 |
不能 |
不能 |
甲基化作用的位点 |
寄主特异性的位点 |
寄主特异性的位点 |
寄主特异性的位点 |
序列特异性切割 |
不是 |
是 |
是 |
在DNA克隆中的用途 |
无用 |
十分有用 |
有用 |
二.Ⅱ型限制要酶的基本性质
3种类型的限制酶中,Ⅱ型酶的相对分子质量范围是2.0×104~1.0×105。它们的核酸内切酶活性和甲基化作用活性是分开的,且只需要Mg2+来激活,并不需要ATP辅助因子;在识别序列和切割位点上,也有其特异性,因而Ⅱ型酶是DNA操作的重要工具酶。通常说的限制酶就是指的Ⅱ型酶。限制酶的反应温度一般为37℃。Ⅱ型限制酶有
3个基本特性:一是在 DNA双链分子上有特异性识别并易切割的序列部位,使被切割的DNA分子形成单链的断裂;二是在DNA分子上两个单链断裂的部位通常不是彼此直接相对的;第三,断裂结果所形成的DNA片断常具有互补的单链延伸末端。
一般,Ⅱ型酸能识别由4—6个(或7个)核苷酸所组成的特定序列,这类序列常通称为限制酶的识别序列。由于限制酶是从它们的识别序列内来切割DNA分子,所以识别序列又称为限制酶的切割位点或靶序列。大多数Ⅱ型酶切割位点的一个显著特点是它们具有双重旋转对称的结构或称回文序列(palindrome)。
切割片段末端的特点:一种是形成粘性末端,它们能够通过互补碱基间的配对重新环化。另一种是形成具有平末端的DNA片段,这种片段则不易于环化。

虽然不同限制酶识别不同靶序列,但也有一些来源不同的限制酶识别同样的靶序列,这类酶就特称为同裂酶或同切点酶(isoschizomer),它们发生同样的切割,产生的粘性末端却不一定相同。与同裂酶相对应的一类限制酶称为同尾酶(isocaudamer),它们来源各异,识别的靶序列也各不相同,但都产生相同的粘性末端。它们产生的粘性末端可以彼此连接起来,然而应注意的是它们重组后所形成的序列结构却不能再被原来的任何一种同尾酶所识别。
三.DNA连接酶及其连接作用
DNA连接酶(ligase)是一种能够催化DNA中相邻的3'-OH和5'-磷酸基末端之间形成磷酸二酯键并把两段DNA拼接起来的核酸酶。它最早发现于1957年,是从T4噬菌体中提取的,它是体外构建重组DNA分子必不可少的工具酶。同限制性内切酶一样,连接酶的发现和应用对基因工程的创立和发展有着举足轻重的意义。
DNA连接酶能够封闭DNA双链,并不能连接两条单链DNA分子,而被连接的DNA链必须是双螺旋DNA分子的一部分。事实上,DNA连接酶所封闭的是双螺旋DNA骨架上的切口(nike)而不能封闭制缺口(gap);这也就是指只有当3'-OH和5'-P彼此相邻,而它们又是各自位于互补碱基配对的两个脱氧核苷酸的末端时,DNA连接酶才以能把它们连接成磷酸二酯键。连接过程的这种形式对于正常DNA的合成、损伤DNA的修复和遗传重组中DNA的拼接都是十分必要的。连接酶催化反应的最适温度是37℃。
连接酶把两个具有粘性末端的DNA分子连接在一起的方式称作“粘接”,而使平末端的双链DNA分子彼此连接的方式则称为“端接”。连接酶也用于连接化学合成的衔接物(linker),这有助于体外合成人造基因。
二.转基因动物技术及其应用
(一)转基因动物的概念
借助基因工程技术把外源目的基因导入生殖细胞、胚胎干细胞和早期胚胎,并在受体染色体上稳定整合,使之经过各种发育途径得到能把外源目的基因传给子代的个体,即转基因动物(transgenic
animal)。转入的目的基因称为转基因(transgene),这种转移目的基因的过程称为转基因作用(transgenesis)。关于“转基因”的概念,通常只限于动、植物中经基因工程技术进行的基因转移,因此品种间不论有性或无性杂交获得新基因的途径都不属此范畴。
(二)转基因动物技术
至今主要有3种途径可以产生转基因动物,即显微注射法、反转录病毒法和胚胎干细胞法。反转录病毒转移基因的基本方法在前面已有简介,这里只介绍显微注射法和胚胎干细胞法两种。
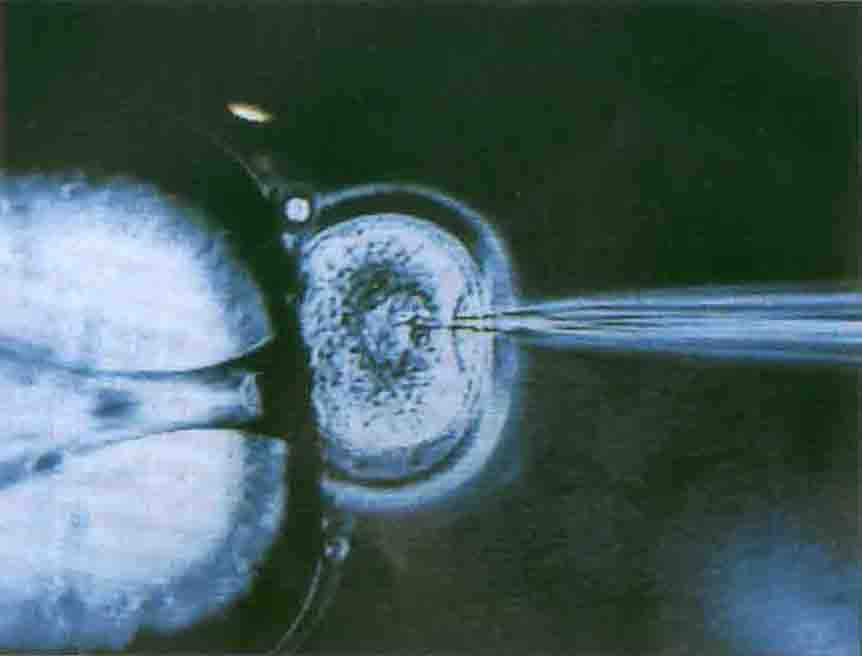
1.受精卵原核显微注射法 显微注射技术是从动物胚胎学研究移核实验的基础上发展而来。已经成功地建立了转基因小鼠、转基因绵羊、转基因猪等等,可见显微注射法是迄今应用得较为普遍而又最有成效的一种获得转基因动物的技术。
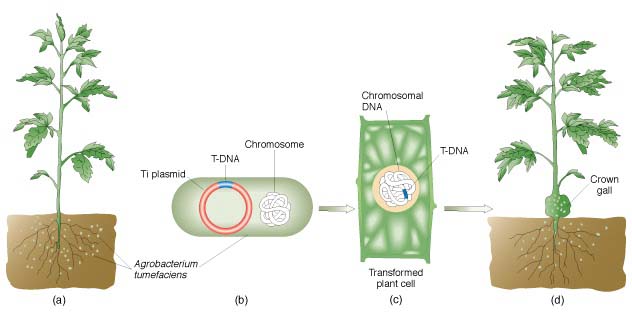
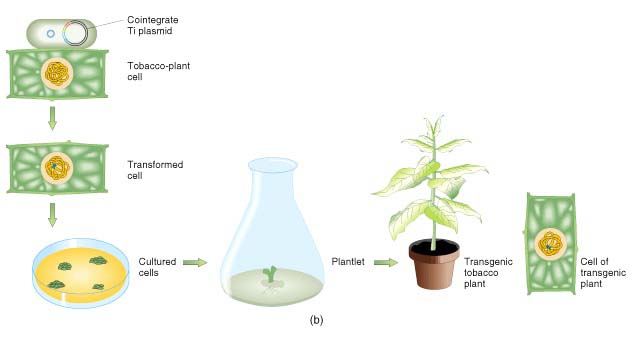
2.胚胎干细胞介导法 胚胎干细胞(embryonic
stem cell,ES)是指从哺乳动物胚胎囊胚期内的细胞团中分离出来的尚未分化的胚胎细胞,这种细胞具有发育的多能性,能够分化出各种组织。这个途径是将外源基因直接导人ES细胞,经体外培养筛选后再注入到受体囊胚腔中,与其中的囊胚细胞聚集在一起,成为受体胚胎的一部分,参与其分化。由这种胚胎发育而成嵌合体的转基因动物,即其中有一部分组织来源于整合有外源基因的供体ES细胞。在嵌合过程中,被转化的ES细胞分化而成的生殖细胞可通过杂交将引人的外源基因传递下去。
3.反转录病毒转移基因 反转录病毒是一类含有RNA的动物病毒,它的基因组进入寄主细胞内,在反转录酶催化下合成DNA。反转录病毒的这种特性说明它有可能是一种天然的转导因子,可经人工操作改建为动物基因的转移载体。另外,反转录病毒感染效率高,但却不会招致寄主细胞的死亡,被它感染或转化的动物细胞常可持续许多世代。反转录病毒可用作载体的另一个优点是其寄主范围广泛,包括无脊椎动物和脊椎动物,其中有的还能够在人体细胞中生长。
(三)转基因动物的应用
转基因动物的研究内容非常广泛,20世纪80~90年代以来从基础理论到应用技术在深度和广度上都有了很大发展,并已日渐由实验室走向生产实践。
1.在基因表达调控研究方面的应用
2.用于基因产品的制备
3.动物品种的培育与改良