 |
第十二章
突变和重组机理
|
| 第一节 突变的分子基础 | |
| 第二节 重组的分子基础 | |
| 第三节 转座遗传因子 | |
| 第四节 DNA损伤的修复 |
|
第十二章
突变和重组机理
|
| 第一节 突变的分子基础 | |
| 第二节 重组的分子基础 | |
| 第三节 转座遗传因子 | |
| 第四节 DNA损伤的修复 |
(二)自发损伤
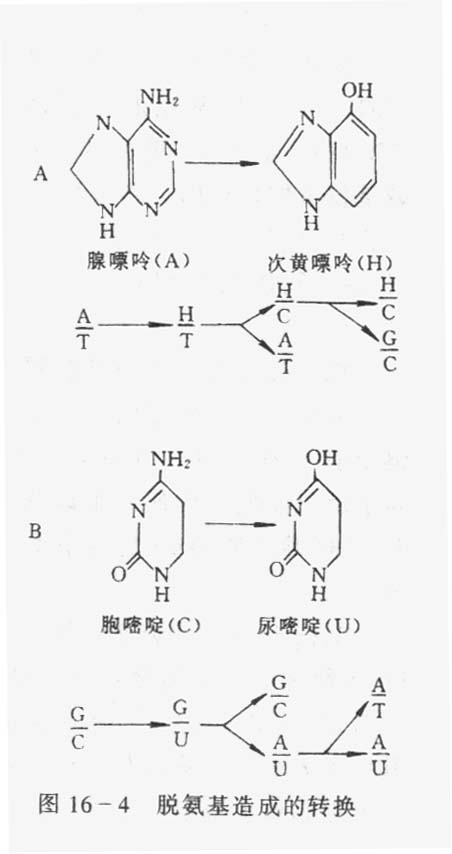
1.脱嘌呤 是由于碱基和脱氧核糖间的糖苷键受到破坏,从而引起一个鸟嘌呤或腺嘌呤从DNA分子上脱落下来。
2.脱氨基 胞嘧啶脱氨基后变成尿嘧啶。
3.氧化性损伤碱基 活泼氧化物如超氧基(O2ˉ),氢氧基(OHˉ)和过氧化氢(H2O2)不仅对DNA的前体,也能对DNA本身造成氧化性损伤,从而引起突变。
二.诱发突变
它是有机体暴露在诱变剂中引起的遗传物质改变,通常比自发突变的频率要高。
(一)诱变机制
诱变剂至少可以通过下列3种机制来诱发突变,它们可以取代DNA中的一个碱基;改变一个碱基使之发生错配;或破坏一个碱基使之在正常情况下无法和任何碱基配对。
1.碱基类似物 某些化学物质和正常的含氮碱基在结构上非常相似,有时它们会替代正常碱基而掺入DNA分子,一旦这些碱基类似物进入DNA分子后,由于它们的配对能力不同于正常碱基,便引起DNA复制过程中其对应位置上插入不正确碱基。
2.特异性错配 某些诱变剂并不掺入DNA,而是通过改变碱基的结构从而引起特异性错配,如某些烷化剂。
一.基因重组的的类型
根据对 DNA序列和所需蛋白质因子的要求,可有3种类型的重组。其共同点是双股DNA间的物质交换,但其发生的情况有所不同。
(一)普遍性或同源重组
它的发生是依赖大范围的DNA同源序列的联会。重组过程中,两个染色体或DNA分子相互交换对等的部分。在真核生物中,重组发生在减数分裂时期的同源染色体的非姊妹染色单体之间。细菌及其某些低等真核生物的转化、细菌的转导、接合以及某些病毒的重组等均属于这一类型。同源重组(homologous
recombination)负责DNA配对和重组的蛋白质因子无碱基序列的特异性,只要两条DNA序列相同或接近,重组就可以在此序列中的任何一点发生。同源重组要求两个DNA分子的序列同源,同源区越长越有利,同源区太短,越难于发生重组。
(二)位点专一性重组(site-specific recombination)
这类重组在原核生物中最为典型。它依赖于小范围同源序列的联会,重组也只限于在这一小范围内,其重组事件只涉及特定位置的短同源区或是特定的碱基序列之间。重组时发生精确的切割、连接反应,DNA不失去,不合成。两个DNA分子并不交换对等的部分,有时是一个DNA分子整合到另一个DNA分子中,因此将这种形式的重组又称整合式重组(integrative
recombination)。
(三)异常重组
异常重组(illegitimate recombination)完全不依赖于序列间的同源性而使一段DNA序列插入到另一段中。但在形成重组分子时往往是依赖于DNA复制而完成重组过程,因此又称为复制性重组(replicative
recombination)。如转座子
二.重组的分子基础
(一)基因转变
重组通常总是交互的。例如在一个杂合体中,如果一染色体把基因A交给它的同源染色体,则它的同源染色体必定把基因a回过来交给它,所以在真菌中,一个座位上的两等位基因分离时,应该呈现2:2分离。可是Lindegren在面包酵母(Saccharomyces
cerevisiae)中发现,有的子囊含有(3A+la)或(1A + 3a)的子囊孢子,而不是预期的(2A+2a)。以后Mitchell在链孢霉的研究中,也发现这个现象,并进行了详细的分析。
Mitchell的杂交试验中,所用的基因是关于吡哆醇(pyridoxine,维生素B6)的合成。在一个位点上,有一突变基因,有这基因的突变株要在培养基上添加吡哆醇后才能生长,但对酸度(pH)是敏感的,改变酸度后,就不要添加了。这突变基因称作pdxp。在一邻近的位点上,也有一突变基因,也是吡哆醇需要型突变,但对酸度是不敏感的。这突变基因称作PdX。这两突变位点非常接近,有迹象表明,它们可能属于同一顺反子。
在一个实验中,Mitchell把两个吡哆醇突变株杂交,+pdxp×pdx+,取得子囊后,对585个子囊中的抱子依次解剖出来,进行培养和鉴定。他发现4个子囊中,有野生型的孢子对(spore
pairs),好象这两位点间有了重组(表12―2);可是跟预期相反,重组后应该同时出现的双突变型(pdx pdxp)孢子对却没有发现。尽管分析方法是灵敏的,如果有双突变型的话,是能够检出的。还有这些情况也不能用突变来说明,因为它们的频率比这些位点的正常突变率高得很多。
上面这些不寻常的情况,好象是由于一个基因转变为另一等位基因,所以称为基因转变(gene conversion)。基因转变往往伴有转换区外基因的重组,但区外基因的重组是正常的交互方式,所以虽然pdxp位点出现异常的3:1(或6:2)分离,但邻接的pdx位点仍显示出正常的2:2分离。同样,基因转变也在粪生粪壳菌(Sordaria
fimicola)中发现。
(二)遗传重组的分子基础
重组不仅有正常的交互方式,而且偶而也有不规则的非交互方式就是所谓基因转变。基因转变往往跟正常互换有关,两者有相当多的机会影响到相同的两条非姊妹单体。还有,有的子囊可以显示减数后分离,表明重组可以影响染色单体,也可影响半染色单体。为了能够说明上面这些事实,一些学者提出不同的重组模型,其中Holliday提出的一个重组模型受到多数学者的支持,以后又经办些作者和
Holliday本人的修改。这个模型叫做异源或杂种DNA模型(heteroduplex or hybrid
DNA model),适用于原核类和真核类。
过去曾经认为,DNA受到损害,或含有不相称的碱基对时,细胞就不能分裂,或产生突变型子细胞。可是近年来研究表明,细胞有时能够修复它的DNA的变异,因为受损害的遗传信息可以从未受伤的互补链获得。
一.紫外线照射对DNA的损伤
紫外线(ultraviolet light, UV)是一种有效的杀菌剂。用紫外线照射细菌,并把细菌培养在黑暗中,那末细菌被杀死的数目与照射剂量成正比。如果细菌接触可见光,大部分细菌就能活下来,这是光能修复辐射引起的损伤的证据。
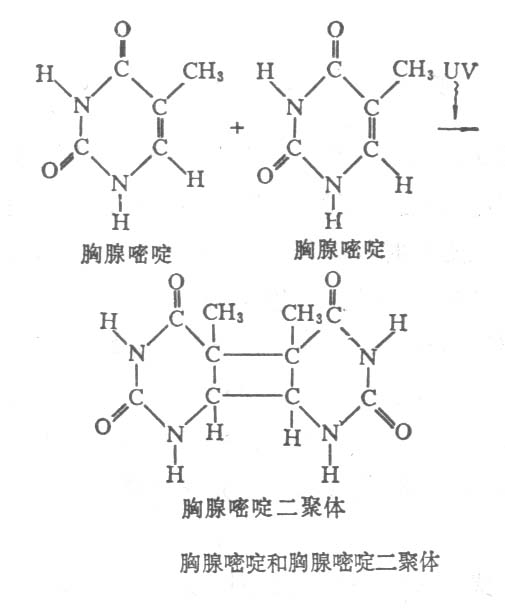
分析紫外线照射后的DNA,最明显的变化是,同一链上的两个邻接嘧啶核苷酸的共价联结,形成嘧啶二聚体。嘧啶二聚体中,最常见的是胸腺嘧啶二聚体(thymine
dimer,TT)(图12-23),此外还有胞嘧啶二聚体(CC)以及胸腺嘧啶和胞嘧啶二聚体(CT)。
这些嘧啶二聚体使双螺旋的两链间的键减弱,使DNA结构局都变形,严重影响照射后DNA的复制和转录。含有嘧啶二聚体的DNA链,使它不能作为DNA复制的样板,新合成的链在二聚体的对面和两旁留下了缺口。
紫外线引起的DNA损伤的修复,大致上通过3个途径:(1)在损伤部位就地修复――光复活(photoreactivation);(2)取代损伤部位――暗修复或切除修复(excision
repair);(3)越过损伤部位而进行修复――重组修复(recombination repair)。
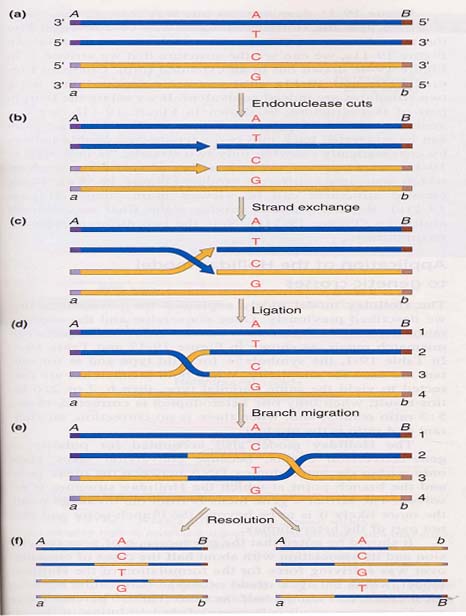
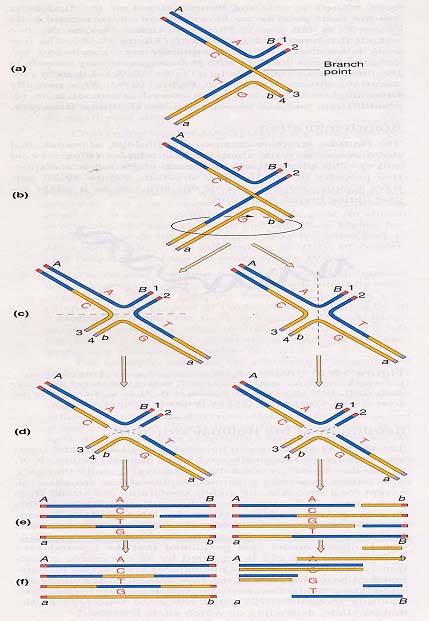
根据Holliday模型,重组过程是这样的:(左图)a.同源的非姊妹染色单体联会;b.姊妹染色单体DNA中两个方向相同的单链(半螺旋),在DNA内切酶作用下,在相同位置上同时切开;c.切开的单链交换重接;d.形成交联桥结构(cross-bridge
structure);e.交联桥的位置可以靠拉练式活动沿着配对DNA分子“移动”。其中互补碱基间形成的氢键从一条亲本链改为另一条亲本链,于是在两个亲本DNA分子间造成一段异源双链DNA,这种结构又称为Holliday结构。(右图)a.左e和右a结构相同;b.绕交联桥旋转180°;c.形成Holliday结构的异构体;d.通过两种方式之一切断DNA单链,恢复两个线性DNA分子;J和K结构相同,如左右切断形成的线性分子是非重组体(Ab、ab),如上下切断形成的线性分子其异源双链DNA区的两侧标记基因是重组体(Ab、aB)。由此可见,不管Holliday结构断裂是否导致旁侧遗传标记的重组,它们都含有一个异源双链DNA区(由G-C、A-T配对变为G-A、C-T非配对)。
3.嵌合剂的致突变作用 嵌入染料,如亚丁晟、原黄素、黄素等亚丁类染料,含有亚丁环,其分子大小与碱基对大小差不多,可以嵌入到DNA双链核心堆积的碱基对之间,在嵌入的位置上能引起单个碱基对的插入或缺失突变;也能嵌入单链DNA的碱基对之间,造成移码突变。
4.辐射的诱变作用
(1)紫外线的致突变作用 紫外线能使DNA产生很多光生成物,其中胸腺嘧啶二聚体是重要的一种损伤。另外,还能引起缺失、重复和移码突变。
(2)电离辐射的诱变作用
5.黄曲霉素B1 这是一种很强的制癌剂。它在鸟嘌呤N-7位置上形成一个加成复合物进而产生无嘌呤位点。
(二)碱基替换对遗传信息的影响
1.同义突变 指不改变氨基酸的密码子变化,这显然与密码子的简并性有关。它不改变氨基酸序列,没有突变效应。
2.错义突变 指碱基替换的结果引起氨基酸序列的 改变。有些错义突变严重影响蛋白质的活性,甚至使活性完全丧失,影响表型。有些错义突变不影响或基本不影响蛋白质活性,不表现明显的性状变化。
3.无义突变 编码区的单碱基突变导致终止密码子(ATG、TGA或TAA)的形成,使mRNA的翻译提前终止,形成不完全的肽链,因而其产物一般是没有活性的。
总之,由碱基替换引起氨基酸序列的改变,将导致蛋白质活性和功能的不同程度的丧失。
(三)移码突变
由于在DNA分子的外显子中插入或缺失1、2或4个核苷酸而导致的阅读框架位移。如果插入或缺失的碱基正好是3个,那么在翻译出的多肽上可能只是多1个或少1个氨基酸,而不完全打乱整个氨基酸序列。
(四)突变热点和增变基因
突变位点在基因内的分布并不是随机的,许多位点上没有突变型或突变型很少,而在某些位点上突变型很多,其突变率大大高于平均数。这些位点称为突变热点。其形成的主要原因是5-甲基胞嘧啶的存在。
增变基因是指基因组中某些基因的突变可使整个基因组的突变率明显上升。主要有两类:DNA多聚酶的各个基因和dam基因。
三.定点诱变
利用人工合成的寡聚核苷酸在离体条件下制造任何部位的位点特异性的技术通常称为定点突变或称定点诱变。在蛋白质工程和基因的表达和调控机制的研究方面具有重要意义。
异源 DNA是不稳定的,不配对的核苷酸在 DNA分子中造成歪斜,不配对的一小段由核酸外切酶(exonuclease)切除,留下单链的缺口。然后在DNA多聚酶的作用下,合成具有互补碱基的区段,填补缺口,再由连接酶的作用,把新合成的短链以共价键联络上去,成为连续的核苷酸链,完成修复过程。
但是在修复时,由于切除的不配对碱基区段的不同,可以有两种方式,如不相称的碱基对G―A的修复,由于切去的区段的不同,或者在染色单体中形成一个野生型基因(+),或者在染色单体中形成一个突变型基因(g)。
如果不相称的核苷酸没有得到校正,杂种DNA(杂种染色单体)留到下一次复制时,将产生两个不同的子染色单体(daughter Chromatids),这样就出现半染色单体转变(half-chromatid
version) ,一个孢子对中的两个孢子不同。
这个杂种 DNA模型清楚地说明,重组是一个酶促过程,不仅DNA多聚酶是合成新的互补链所必需的,而且核酸内切酶,外切酶和 连接酶等也是DNA链的切割,断链的切除和断链的愈合所必需的。
这个模型说明了遗传重组的过程,同时也解释了基因转变的现象,有一定的实验证据,是比较完善的模型。但是随着科学研究的进展,这个模型将会有或多或少的修改,甚而重新建立。那也是可能的。


(一)光复活
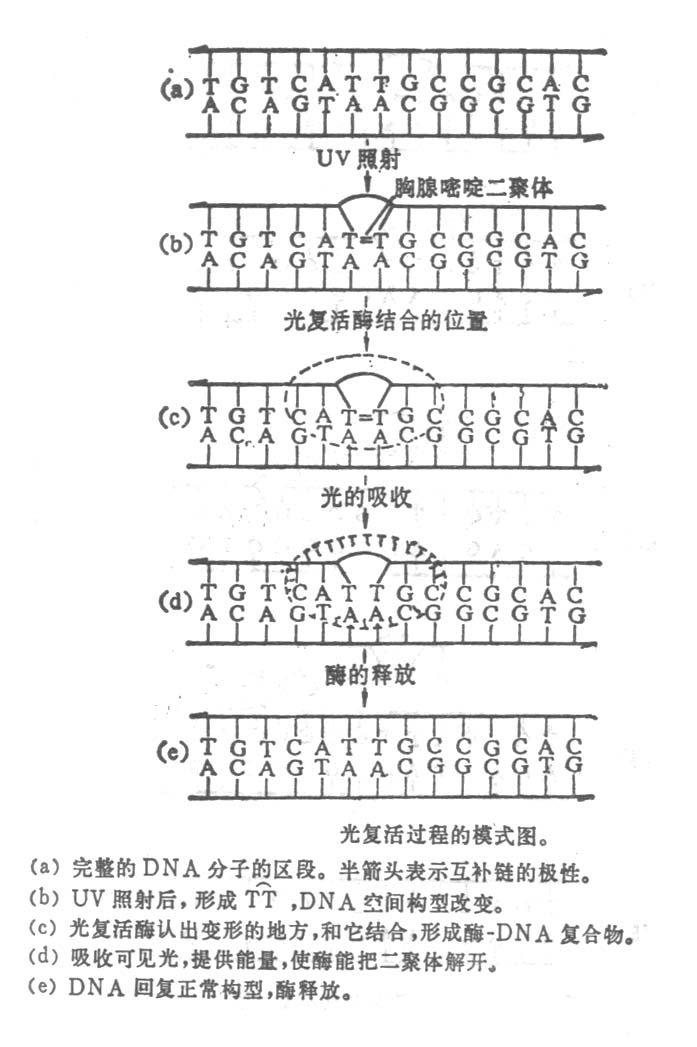
细菌经紫外线照射后,再放在波长310~440nm的可见灯,存活率大大提高,并且降低了突变频率。这是什么原因呢?后来发现这个效应是一种光复活酶(photobreacting
enzyme)的作用。在暗处,光复活酶能认出紫外线照射所形成的嘧啶二聚体,如TT,并和它结合,形成酶和DNA的复合物,颇能解开二聚体。但照以可见光时,这酶利用可见光提供的能量,使二聚体解开成为单体,然后酶从复合物中释放出来,修复过程完成。
光复活酶已在许多生物体内发现,虽然这种修复功能普遍存在,但主要是低等生物的一种修复形式,随着生物进化地位的上升,它所起的作用随之削弱。
(二)暗复活
暗复活过程具有更重要的意义,它并不表示修复过程在黑暗中进行,而只是说,光不起任何作用。这种修复过程不是简单地由另一种酶来拆开二聚体,而是利用双链DNA中一段完整的互补链,去恢复损伤链所丧失的信息;就是把含有二聚体的DNA片段切除,然后通过新的核苷酸链的再合成进行修补,所以又叫做切除修复。切除修复有两种情况,一是先补后切,一是先切后补。一般认为先补后切比较合理。切除修复不仅能除去嘧啶二聚体,而且还可以除去DNA上其它的损害。
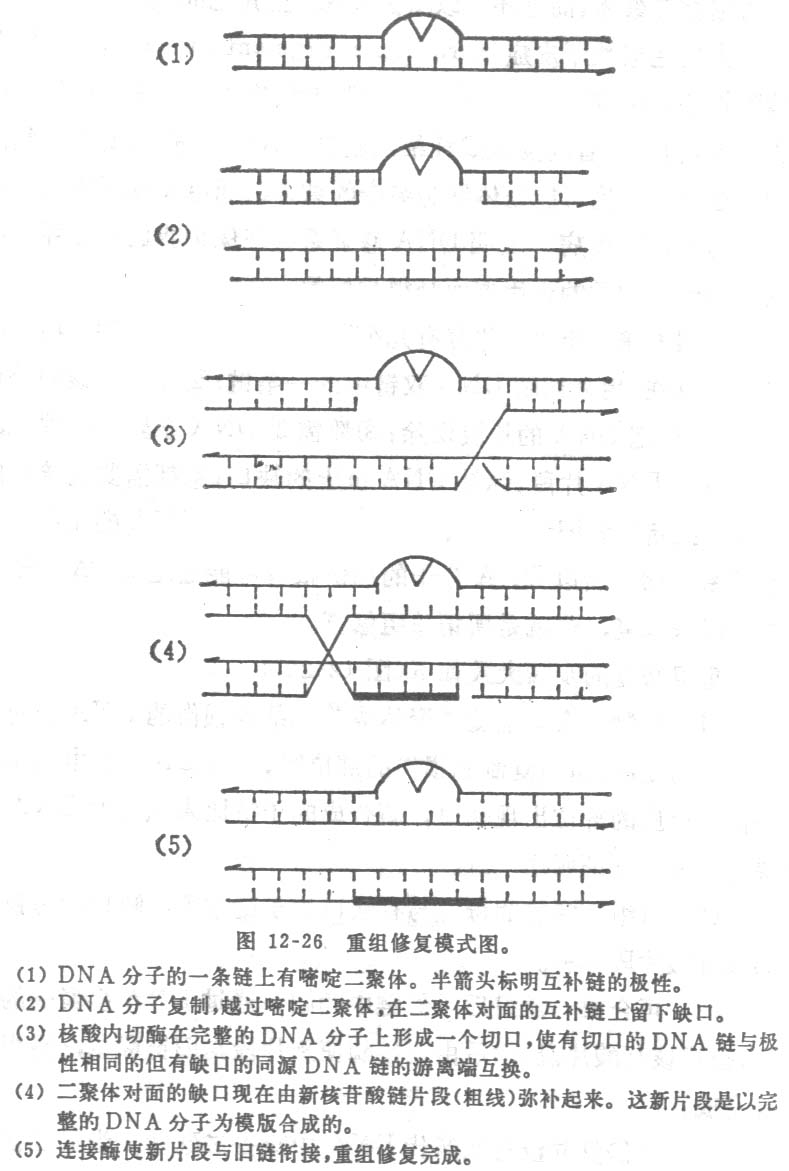
(三)重组修复
重组和修复有几个共同的地方:①都需要核酸内切酶的存在,用来切断 DNA双链中的一条链;②都需要核酸外切酶的参与,把 DNA的片段切除;③都需要DNA多聚酶的催化,合成单链DNA片段,弥补DNA链上的缺口;④都需要连接酶的作用,把新链和旧链以共价键连接起来。因为DNA的重组和修复关系密切,所以DNA分子的损伤很有可能通过DNA分子间的重组来修复,这就是所谓重组修复。
重组修复的步骤大致如下:
(1)复制 含有嘧啶二聚体或其它结构损伤的DNA仍可进行正常的复制。但当复制到损伤的部位时,子代DNA链中与损伤部位相对应的部位出现缺口,新合成的子链比未损伤的DNA链要短一些,就证明这一点。
(2)重组 完整的母链与有缺口的子链重组,缺口由母链来的核苷酸片段弥补。
(3)再合成 重组后,母链中的缺口通过DNA多聚酶的作用,合成核甘酸片段,然后由连接酶使新片段与旧链联结,重组修复完成。
但重组修复并没有从亲代DNA中除去二聚体。当第二次复制时,留在母链中的二聚体仍使复制不能正常进行,复制经过损伤的部位时所产生的缺口,仍旧要用同样的重组过程来弥补。随着复制的继续,若干代以后,虽然二聚体始终没有除去,但损伤的DNA链逐渐“稀释”,最后终于无损于正常的生理过程,损伤也就得到了修复。
二.电离辐射引起的DNA损伤和它的修复
X射线的作用不象紫外线那样有选择性。除射线的直接作用外,还通过水的电离所形成的自由基起作用(间接作用)。DNA链可以产生双链断裂,也可以产生单链断裂,造成缺失、重复、倒位和易位。高剂量照射时,还有碱基的破坏。因此电离辐射的作用比较复杂,修复机理还远远没有紫外线清楚。不过随着电离辐射在育种,临床等方面的应用,电离辐射损伤的修复机理的研究也越来越多。
近几年来,采用E.coli K12 的各种变异株,研究了X射线损伤的修复。实验结果认为,在各种情况下,X射线引起的原初 DNA单链断裂相同,只是由于修复效应的差异,所以观察到的单链打断有差别,未被修复的单链断裂可以是致死的损伤。通常认为可分3种修复过程,需要3种不同的修复酶系。
(1)超快修复 单链打断的极快修复过程。0℃时,两分钟内即可完成,可能是 DNA连接酶的单独作用。E.coli在缺氧条件下照射时,观察到的单链打断少于有氧条件下的照射。
(2)快修复 需要DNA多聚酶 I的修复过程,室温下在缓冲溶液中迅速进行。照射后,细菌在室温下缓冲液中放置几分钟,超快修复后剩下的单链断裂有
90%可被修复。缺乏DNA多聚酶I的 E.coli变异株经X线照射后,可以观察到较多的单链断裂,这是由于缺乏这种修复系统的结果。但是它与紫外线损伤的切除修复不完全相同,在快修复过程中,不需要对嘧啶二聚体专一作用的核酸内切酶。
(3)慢修复 细菌照射后,在37℃培养基中培养40―60分钟,快修复所不能修复的单链断裂,可由重组修复系统修复。因而当重组功能发生障碍时,细菌对X射线的敏感性显著增加。
修复过程在生物体内是普遍存在的,也是正常的生理过程。不仅紫外线和电离辐射引起的损伤可以被修复,许多化学诱变剂,从亚硝酸那样的简单化合物到氮芥,丝裂霉素C等复杂的致癌物质所引起的损伤也能被修复。对紫外线敏感的细菌,往往对电高辐射和化学诱变剂也敏感,抗紫外线辐射的菌株对其他诱变因素也往往有抗性。
越来越多的证据表明,原核生物和真核生物的细胞中存在着多种不同的修复途径。在漫长的进化过程中,选择有利于修复过程的多样化和完善化,但遗传信息的传递过程仍然不可能是完美无缺的,事实上,DNA顺序的改变时时以低频率发生着,为进化过程中自然选择提供新的原材料,使各类生物能在选择作用下不断地向前发展。