 |
第十一章遗传的分子基础 |
| 第一节 遗传物质是DNA(或RNA) | |
| 第二节 DNA的分子结构 | |
| 第三节 DNA与蛋白质 | |
| 第四节 基因的近代概念 |
|
第十一章遗传的分子基础 |
| 第一节 遗传物质是DNA(或RNA) | |
| 第二节 DNA的分子结构 | |
| 第三节 DNA与蛋白质 | |
| 第四节 基因的近代概念 |
第一节 遗传物质是DNA(或RNA)
染色体是由DNA和两类蛋白质:组蛋白(histones)和非组蛋白(non-histones),以及RNA构成。其主要成分是DNA和组蛋白,虽然这两种成分都在基因功能上起着重要的作用,但基因的主要特性由DNA决定,或者说遗传信息贮存在DNA中。
一.DNA是遗传物质的间接证据
(1)DNA通常只在核中的染色体上找到。
(2)同一种生物,不论年龄大小,不论身体的哪一种组织,在一定条件下,每个细胞核的DNA含量基本是相同的,而精子的DNA含量正好是体细胞的一半。蛋白质等其它化学物质不符合这种情况。
(3)同一种生物的各种细胞中,DNA在量上恒定,在质上也恒定;相反,蛋白质在量上和质上都不恒定。
(4)各类生物中,能改变DNA结构的化学物质都可以引起突变。
二.DNA是遗传物质的直接证据
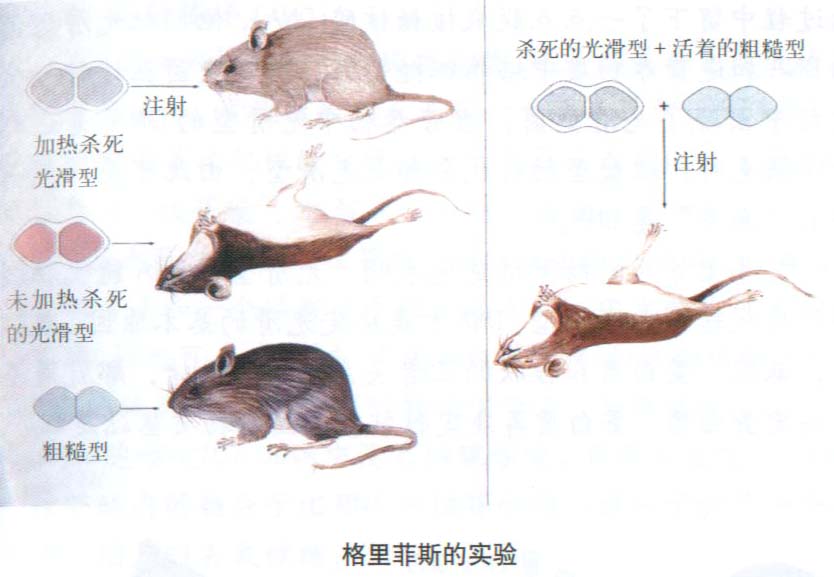
(1) 肺炎球菌的转化实验
肺炎球菌(Diplococcus pneumonoae)有许多不同的菌株(strains),但只有光滑型(S)菌株能引起人的肺炎和小鼠的败血症(septicemia)。当这种细菌生长在合成培养基上时,每个细菌长成一个明亮、光滑的菌落。其他一些菌株没有荚膜,不会引起疾病,长成粗糙型(R)菌落。英国卫生部病理学实验室的Fred
Griffith发现(1928),将高温杀死的S型细菌和活的R型细菌一起注入小鼠体内,结果不仅有许多小鼠死于败血症,而且从死鼠血液中还发现了活的S型细菌。可是,如果注入小鼠体内的只是活的R型细菌,或是死的S型细菌,都不会引起败血症。这说明,高温杀死的S型细菌使某些活的R型细菌转化成S型细菌。S型细菌有一种物质或转化因素(transforming
principle)进入了R型细菌,引起R型细菌发生了稳定的遗传变异。艾弗里(Osward Avery)等人(1944)从S型细菌中分别抽提出DNA、蛋白质和荚膜物质,并把每一种成分同活的R型细菌混合,悬浮在合成培养液中。结果发现只有DNA组分能够把R型细菌转变成S型细菌。而且DNA的纯度越高,这种转化的效率也越高。这说明,一种基因型细胞的DNA进人另一种基因型的细胞后,可引起稳定的遗传变异,DNA赋有特定的遗传特性。
(3)
烟草花叶病毒的重建实验
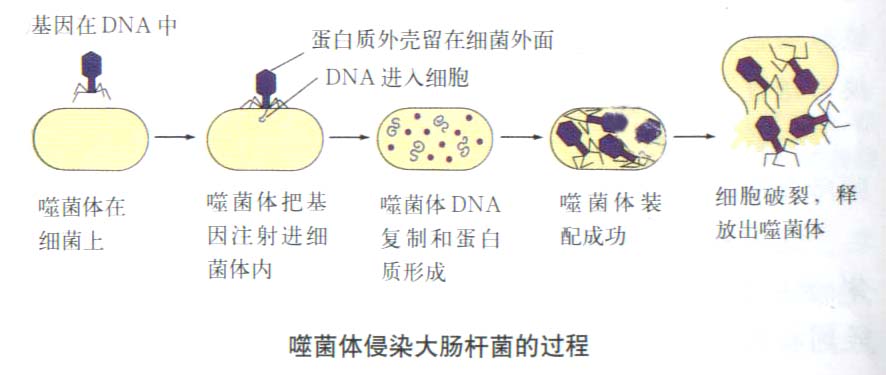
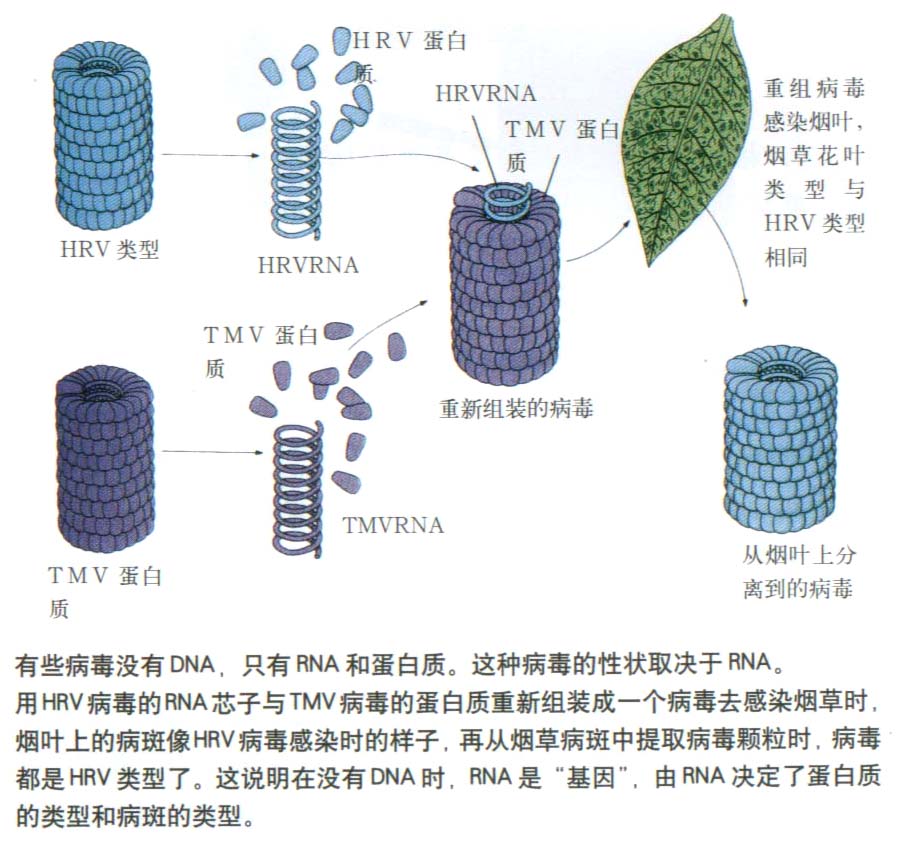
对病毒的研究逐渐深入以后,发现有很多种病毒不含DNA,而是由核糖核酸(RNA)和蛋白质组成。用RNA病毒做的病毒重建实验证明,在只有 RNA而不含DNA的病毒中,遗传物质是RNA。
Fraenkel Conrat(1956)用一种RNA病毒――烟草花叶病毒(tobacco mosaic virus, TMV)作为实验材料。TMV有一个圆筒状蛋白质外壳,由很多个相同的蛋白质亚基组成。外壳内有一条单链的RNA分子沿着其内壁在蛋白质亚基间盘旋着。TMV中94%是蛋白质,6%是RNA。现在要证明在这种RNA病毒中的遗传物质是RNA而不是蛋白质。
把TMV在水和苯酚中震荡,使TMV的蛋白质和RNA分开,然后分别用来感染烟草。只用TMV的蛋白质无法感染烟草;可是,只用TMV的RNA就能感染烟草。病毒RNA进入烟叶细胞后进行复制增殖,生成正常的病毒子代。不过单用RNA感染时效率很低,这可能是因为在感染过程中,病毒RNA被宿主细胞内的酶破坏了一部分。
TMV有许多株系,它们可感染不同的宿主植株,并可在宿主植株的叶片上形成不同类型的病斑。例如,有不同外壳蛋白质的两个株系,S株系的外壳蛋白质不含组氨酸和甲硫氨酸,HR株系的外壳蛋白质则含有这两种氨基酸。Fraenkel
Conrat先将病毒的蛋白质外壳同RNA分开,然后把S株系的蛋白质外壳同HR株系的RNA组合形成杂种病毒。这些有着S株系蛋白质外壳的杂种病毒可被抗S株系的抗体所灭活,但不受抗HR株系的抗体的作用。杂种病毒感染烟草叶片后,形成的病斑类型与RNA供体(即HR株系)的病斑相同,而且从病斑中分离出来的病毒经抗HR株系的抗体处理后也就失去了感染活性。这说明,杂种病毒产生的第二代病毒既含有HR株系的RNA,而且也具有HR株系的蛋白质外壳。当把HR株系的蛋白质外壳同S株系的RNA组合重建成杂种病毒,用来感染烟草时,病斑的类型和第二代病毒的蛋白质外壳也是同RNA供体的株系一样。简言之,当用A的蛋白质与B的RNA,或用A的RNA与B的蛋白质重建成杂种病毒时,决定杂种病毒遗传性状的是RNA,RNA是遗传物质。
综上所述,在DNA的生物中,DNA是遗传物质,在不含DNA而只含有RNA的病毒中,RNA是遗传物质。
一.DNA的组成成分
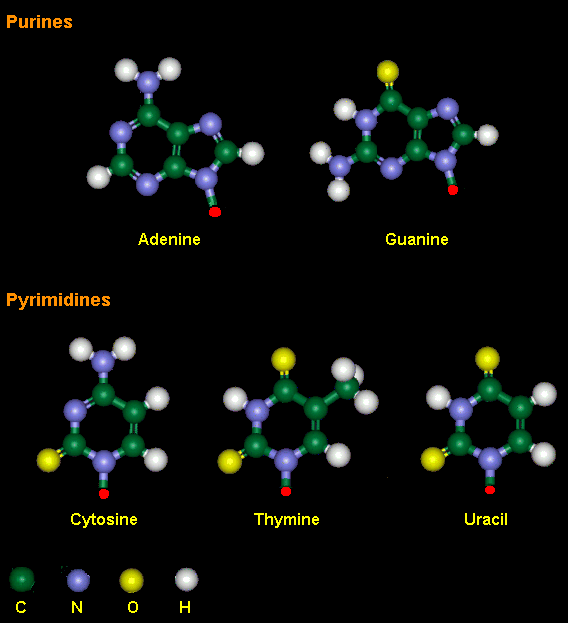
DNA是一种化学分子,基本的组成成分是碱基、脱氧核糖和磷酸。碱基(base)分4种,其中2种是嘌呤,是腺嘌呤(adenine,A)和鸟嘌吟(guanine,G);另外2种是嘧啶,即胞嘧啶(cytosine,C)和胸腺嘧啶(thymidine,T)。碱基与脱氧核糖结合,构成脱氧核糖核苷(deoxyribonucleoside)。脱氧核糖核苷分子中的糖分子与磷酸分子结合,构成脱氧核糖核苷酸(deoxyribonucleotide),核苷酸依据其组成中的碱基种类不同,分别称为腺嘌呤核苷酸(A)、鸟嘌呤核苷酸(G)、胞嘧啶核苷酸(C)和胸腺嘧啶核苷酸(T)。每个核苷酸通过磷酸和糖连接在一起形成了糖-磷酸-糖-磷酸的长链,构成了DNA分子的“脊梁骨”。
RNA的组成与DNA的组成有两点不同:①RNA的4种碱基中,没有胸腺嘧啶(T)而代之以尿嘧啶(uracil,U);②RNA含有的五碳糖是核糖而不是脱氧核糖。
美国哥伦比亚大学的查加夫(Edwin Chargaff)定量分析了 DNA的碱基组成。1951年他指出,在他研究过的物种的DNA碱基组成中,腺嘌呤(A)的量总是几乎与胸腺嘧啶(T)相等;鸟嘌呤(G)则与胞嘧啶(C)几乎相等。在数量上A=T,G=C的关系被称为查加夫法则(Chargaff’s
rule),这对弄清DNA的结构起了重要作用。
二.DNA的结构
1931年英国科学家阿斯特伯里用瑞典科学家从胸腺制备的高相对分子质量DNA作材料,首次得到了其X光衍射图像。
1950英国物理学家威尔金斯(Maurice Wilkins)用真正的DNA晶体做X光衍射分析,确定DNA有很精细的结构。每条DNA链尽管各有自己特有的核苷酸组成和排列,但彼此被连接成十分相似的结构。
接着,威尔金斯的同事弗兰克琳(Rosalind Franklin)用在很高湿度下含水较多的DNA做X光衍射。她从得到的图像中判定DNA是螺旋结构。在DNA长链上,每个核苷酸相距0.34nm,每一个螺距为3.4nm,有10个核苷酸。以前测定DNA的宽度为2nm,这个数据是十分重要的。因为如果DNA分子只含一条链,那么2nm的宽度就太大了。因此推测,DNA分子是由不止一条链所构成的。
1953年英国《自然》杂志发表了时年仅25岁的沃森(Watson J D)和35岁的克里克(Crick
F H C)的研究论文《核酸的分子结构――脱氧核糖核酸的结构》,刊出了DNA的双螺旋结构模型,从而正确地解析了DNA的结构和功能之间的关系,由此使遗传学进入了一个新天地。沃森和克里克制作的DNA双螺旋结构模型与X光衍射提供的螺旋构型的参数相吻合。

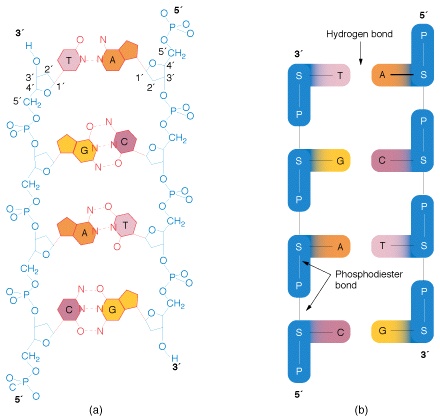
DNA分子的糖-磷酸组成的“脊梁骨”位于DNA的外侧,嘌呤和嘧啶碱基位于DNA的内侧。一条链上的碱基通过氢键与另一条链上的碱基配对连接。两条链上的碱基之间的配对并非是随机的,而总是A与T通过形成的2个氢键相连接(A=T或T=A),C与G则通过形成的3个氢键而配对(C≡G或G≡C)。这样,沿着任何一条DNA链,所有4种碱基有着多种多样可能的排列顺序。
两条通过碱基间配对连接的DNA链,称为互补链(complementary chain)两条互补链呈反向平行的螺旋结构,每一螺距为3.4nm,内有10个碱基对。尽管形成碱基间配对连接的氢键是非常弱的键,但由于互补链之间有着这么多的碱基对,所以在生理条件下从来不会自发地分开。可是,如果在高温下,DNA分子的氢键就会遭到破坏,于是双螺旋解离成两条互补链。这一过程称为变性(denaturation)。变性后的互补链再回到正常条件下,又会重新连接成双螺旋结构。这一过程称为复性(renaturation)。由于AT之间的氢键有2个,CC之间有3个氢键,所以就变性和复性而言,AT比GC容易。
三.DNA双螺旋结构的生物学意义
DNA双螺旋结构可以很完美地解释基因的复制和传递。DNA双螺旋是由两条互补链组成的,同时碱基之间的配对是有规律的。因此,当DNA双链分开成为两条互补链单键时,单链可以作为复制的模板,再重新合成与它互补的DNA链。这样,原来的DNA双链分子,也就是亲链(perental chain),通过复制产生了两个DNA双链分子,也就是两个子链。子链中的碱基对同亲链完全一样。DNA分子就是这样复制出与自己完全相同的两个子代。
生命活动是通过蛋白质来实现的。不同的生物有不同的结构和性状,且是遗传的;也就是说,这是由基因决定的。因此,基因决定生物的遗传性状是通过蛋白质来实现的。基因将所携带的遗传信息传递给蛋白质。不同的基因决定不同的蛋白质。
一.蛋白质的组分与结构
蛋白质是由氨基酸组成的肽链折叠而成的大分子物质。蛋白质中常见的氨基酸有20种。氨基酸有着共同的骨架,不同的氨基酸则有不同的侧链(简称R基)。一个氨基酸的羧基与另一个氨基酸的氨基通过肽键(―CO―NH―)连接,反应过程中失去一分子水(H2O)。氨基酸以这种方式连接成一条多肽链。少于20个氨基酸连接成的短链称为肽(peptide)。两个氨基酸连接成的称为二肽,三个氨基酸连接成的称为三肽,依此类推。
多肽链在三维空间是折叠的。一种是形成α螺旋。在这种结构中,每一圈螺旋含有3.6个氨基酸残基。在α螺旋里,羧基氧处于某一适当的位置,与相隔3个残基的氮上面的氢原子形成氢键。另一种结构是肽链的β片层结构。在这种结构里,肽链按相反方向伸展并通过氢键形成片层状的折叠。
蛋白质分子折叠的三维空间构型同多肽链中氨基酸的组成和排列顺序有关。除氨基酸之间形成的氢键外,侧链也能够牵制一部分主链,有助于改变主链的方向,通常能使蛋白质肽链产生一个转角。含硫的半胱氨酸的侧链上能形成增加蛋白质空间结构的稳定性的二硫键。除此之外,蛋白质三维结构中疏水的氨基酸残基深藏于蛋白质分子的内部,亲水的氨基酸残基则暴露于蛋白质分子与水接触的表面。肽链的折叠除了取决于肽链中氨基酸的组成和排列顺序外,还有赖于称为分子陪伴的一类分子。
蛋白质的结构可以分成等级:
一级结构:肽链中的氨基酸的线性排列结构。
二级结构:肽链的α螺旋和β片层结构。
三级结构:一条肽链的三维空间的折叠结构。
四级结构:几条肽链(或几个亚基)结合形成的三维空间的折叠结构。
尽管蛋白质高级结构的形成与一级结构密切相关,可是,不同的一级结构也能决定相同的三级结构。蛋白质是由许多结构域(domain)所组成。一个结构域是蛋白质的一个相对独立的区段,在有些情况下,一个结构域可通过多肽链中的相关片段单独形成自身的构象。变构蛋白质(allosteric
protein)可以有不同的构象(conformation),不同的构象具有不同的生物学性质。构象之间的变换受到蛋白质和辅助因子(cofactor)以及蛋白质与蛋白质之间的相互作用的影响。变构蛋白质在代谢和遗传调控中起重要作用。
二.基因的特性
1.基因能自主复制
2.基因能决定性状
3.基因能发生突变
三.基因的类型
(一)根据基因的功能和性质,可将其分为以下几类:
1.结构基因(structural genes)与调节基因(regulatory
genes) 这类基因不仅可转录(transcroption)成mRNA,而且可翻译(translation)成多肽链,从而构成各种结构蛋白和催化各种生化反应的酶。调节基因的作用是调控其他基因的活性,调节基因可转录成mRNA,然后由mRNA再翻译成阻遏蛋白质或激活蛋白质。
2.核糖体RNA基因 (ribosomal RNA genes,简称rDNA)与转移RNA基因(transfer
RNA genes,简称tDNA) 这类基因只转录产生相应的RNA,而不翻译成多肽链。rDNA是专门转录核糖体RNA(rRNA)的,rRNA与相应的蛋白质结合形成核糖体,为mRNA翻译形成多肽链提供场所;tDNA专门转录转移RNA(tRNA),tRNA的作用是激活氨基酸,因为在多肽链合成时,氨基酸先要被激活,然后被转移到核糖体上按mRNA信息与其他氨基酸连接形成多肽链。显然该类基因与蛋白质合成密切相关,而且都是多拷贝,因此这类基因即使少数拷贝发生变化,一般也不会带来严重后果。
3.启动子(promotor)与操纵基因(operator)
前者是转录时RNA多聚酶起始与DNA结合的部位;后者是调节基因的产物阻遏蛋白质或激活蛋白质与DNA结合的部位,它们都是不转录的DNA区段,确切说,它们不能称为基因。但关系到结构基因的活化或钝化。
(二)重叠基因(overlapping gene)
1978年,英国剑桥分子生物学家Sanger分析发现重叠基因。所谓重叠基因是指两个或两个以上的基因共有一段DNA序列,或是指一段DNA序列成为两个或两个以上基因的组成部分。
(三)断裂基因(split gene)
1977年法国的Chambon等和美国的Berger等首次报道了基因内部有间隔顺序(spacer sequence)。由于基因内部有间隔顺序,从而提出断裂基因的概念就是一个基因由几个互不相邻的段落组成,它们内部被长达数百个乃至上千个核苷酸对的间隔序列所隔开。
DNA是完整的,只是有些片段找不到可以和它配对的mRNA,于是这段DNA就拱起来。显然这些拱起来的DNA序列可能未被转录,至少在成熟的mRNA序列中没有这部分DNA的转录产物。因此将DNA序列中被转录成为mRNA的片段称为外显子(exon或extron),而在成熟mRNA
上未反应出的DNA片段称为内含子(intron)。
(四)基因家族(gene family)和基因簇(gene cluster)
1.基因家族(gene family)
真核生物的基因组中有许多来源相同、结构相似、功能相关的基因,这样的一组基因称为一个基因家族。一个基因家族中的成员可以紧密地集中在一处,中间常以中度重复序列相间隔。但同一基因家族成员可分散在不同的染色体上,也可以集中在一条染色体上。如珠蛋白基因家族,Alu家族,Hinf家族。
根据基因家族的复杂程度,可以把它们区分成几种类型:①简单的多基因家族;②复杂的多基因家族;③不同场合表达的复杂多基因家族。
2.基因簇(gene cluster)
一个基因家族的成员紧密连锁成簇状排列在某一染色体上,形成一个基因簇(gene cluster)。人类基因组中有几个大的基因簇,例如为血红蛋白分子两种多肽编码的α珠蛋白基因簇和β珠蛋白基因簇,编码免疫球蛋白重链和轻链的基因复合体,以及编码人类主要组织相容性复合体(MHC)HLA基因簇等。
在多基因家族中,某些成员并不产生有功能的基因产物,但在结构和DNA序列上与有功能的基因具有相似性,这种成员称为假基因(pseudogene)。假基因与有功能基因同源,原来可能是有功能的基因,但由于缺失,倒位或突变等原因使该基因失去活性成为无功能基因。最早发现的假基因是在非洲爪蟾的5SrRNA基因中。
五.基因组
(一)基因组和C值
一个物种单倍体的染色体的数目及其所携带的全部基因称为该物种的基因组(genome)。一个物种单倍体基因组的DNA含量是相对恒定的,它通常称为该物种DNA的C值,不同物种的C值差异极大,最小的C值是支原体,最大的是某些显花植物和两栖动物的C值。从原核生物到真核生物,其基因组大小和DNA含量是随着生物进化复杂程度的增加而稳步上升的。随着生物结构和功能复杂程度的增加,需要的基因数目和基因产物种类越多,因而C值也越大。
尽管C值大小随着生物的复杂程度增加,然而在结构和功能相似的同一类生物中,以致亲缘关系很近的物种之间,它们的C值差异可达10倍乃至上百倍,如两栖类和被子植物不同物种之间。因此,C值的大小并不能完全说明生物进化的程度和复杂性的高低,也就是说,物种的C值和它进化复杂性之间没有严格的对应关系,这种现象称为C值悖理(C
value paradox)。C值悖理现象说明真核生物基因组中必然存在大量的不编码基因产物的DNA序列。
(二)DNA序列的类别
单拷贝序列(unique sequence) 亦称非重复序列(nonrepetitive
sequence) 在一个基因组只有一个拷贝或2-3个拷贝。
中度重复序列 (moderately sequence) 中度重复序列中的重复单位平均长度约300
bp,重复次数为10-102。如人的珠蛋白(血红蛋白)基因。
高度重复序列(highly sequence) 就是在基因组中存在大量拷贝的序列,一般重复次数在106以上,通常这些序列的长度为6-200bp,如卫星DNA。
卫星DNA(satellite DNA) 是一类高度重复的DNA序列。在氯化铯梯度离心中,在主要DNA带的前面或后面有一个次要的DNA带相伴随,这些小的区带就象卫星一样围绕着DNA主带,故称卫星DNA。
(三)基因组研究
基因组学(genomics)是研究生物体基因和基因组的结构组成、不稳定性及功能的学科。基因组学可分为结构基因组学(structural
genomics)和功能基因组学(functional genomics)。前者是研究基因和基因组的结构,各种遗传元件的序列特征,基因组作图和基因定位等;后者着重研究不同的序列结构具有的不同功能,基因表达的调控,基因和环境之间(包括基因与基因之间,基因与其他DNA序列之间,基因与蛋白质之间)相互作用等。
转录组学(transcriptomics)是研究转录组(transcriptome)的一门学科,即研究某一时刻某一细胞里基因组转录产生的全部转录物的种类、结构和功能。
蛋白质组学(proteomics)是一门研究细胞内全部蛋白质(即蛋白质组,proteome)的组成及其活动规律的学科。
表型组学(phenomics)是研究生物体整个表型(phenotype)形成的机制的学科。
一.基因的概念
(一)遗传“因子” 基因的最初概念是来自孟德尔的遗传“因子”,认为生物性状的遗传是由遗传因子所控制的,性状本身是不能遗传的,控制性状的遗传因子才是遗传的。1909年,丹麦学者W.L.Johannsen提出了“基因”(gene)一词,代替了孟德尔的遗传因子,并由此形成了“颗粒遗传”学说,认为在杂种中等位基因不融合,各自保持其独立性,这也是孟德尔遗传规律的核心。
(二)染色体是基因的载体 1910年摩尔根等通过果蝇杂交实验表明,染色体在细胞分裂时的行为与基因行为一致,从而证明基因位于染色体上,并呈直线排列,提出了遗传学的连锁交换规律,证明了性别决定是受染色体支配的,并于1926年发表了《基因论》。
这一理论揭示了基因与性状之间的关系和遗传的传递规律,建立了遗传的染色体学说,为细胞遗传学奠定了重要基础。他本人1933年获诺贝尔奖金。摩尔根科学地预见了基因是一个化学实体,并认为基因控制相应的性状,基因可以发生突变,基因之间可以发生交换,由此提出基因既是一个功能单位,是一个突变单位,也是一个交换单位的“三位一体”概念。
(三)DNA是遗传物质 自摩尔根以后,几位科学家研究证明多线染色体与生殖细胞染色体之间有对应关系,提出多线染色体上的横纹就是基因的假说,并得到实验证实,从而把形式上的基因推向实体;1928年
Griffith首先发现了肺炎球菌的转化作用,即用高温杀死有致病力的S品系细菌,可改变无致病力的R品系成为有致病力细菌的效应。这种改变遗传性状的现象称为细菌的转化(transformation),该实验为确定遗传物质化学性质的研究开辟了道路。1944年Avery等人不仅在体外成功地重复了上述实验,而且用生物化学方法证明了转化因子(transforming
factor)是DNA,而不是多糖荚膜、蛋白质和RNA,而且转化频率随着DNA纯度的提高而增加。用DNA酶(DNase)处理使DNA降解后,则不出现转化现象,但其他酶,如蛋白酶等对这种转化能力没有影响。同时这种转化作用也可在体外进行。该实验充分地证明了使R型转化为S型的是S型的DNA片段的转入,证明了DNA就是遗传物质。
(四)基因是有功能的DNA片段 20世纪 40年代 G.W.Beadle和 E.L.Tatum通过对粗糙脉孢菌营养缺陷型的研究,提出了一个基因一个酶的假说,这一假设沟通了生物化学中蛋白质合成的研究与遗传学中基因功能的研究。也为遗传密码的解码和细胞内大分子之间信息传递过程的揭示奠定了基础。特别是
1953年 Watson和 Crick提出了DNA双螺旋结构模型,明确了DNA在活体内的复制方式。1957年由Crick最早提出遗传信息在细胞内的生物大分子间转移的基本法则,即中心法则,接着在1961年又提出了三联遗传密码,这样将DNA分子的结构与生物学功能有机地统一起来,也为揭示基因的本质奠定了分子基础。1957年S.Benzer用大肠杆菌T4噬菌体作为材料,在DNA分子结构的水平上,分析了基因内部的精细结构,提出了顺反子(cistor)概念,证明基因是DNA分子上的一个特定的区段,就其功能来说是一个独立的单位。但在这一特定的DNA片段内含有许多突变位点,也称突变子(muton),即突变后可以产生变异的最小单位。这些突变位点之间可以发生重组,因此一个基因内含有多个重组单位,也称重组子(recon),即不能由重组分开的最小单位。从理论上分析,基因内每一对核苷酸的改变就可导致一个突变的发生,每两对核苷酸之间就可发生重组。由此可见,一个基因具有多少对核苷酸就有多少个突变子和相应数目的重组子,但实际上突变子的数目小于核苷酸对数,重组子数小于突变子数。总之,顺反子学说打破了“三位一体”的基因概念,把基因具体化为DNA分子上特定的一段顺序,即负责编码特定的遗传信息的功能单位――顺反子。但其内部又是可分的,包含多个突变和重组单位。
(五)操纵子模型 1961法国分子生物学家 F.Jacob和J.Monod通过不同的大肠杆菌乳糖代谢突变体来研究基因的作用,提出了操纵子模型学说(operon
theory)。这一学说阐明了基因调控在乳糖利用中所起的作用。即把生物活性相关基因组织在一起进行统一的调控,并保持各基因产物的精确比例。原核生物的操纵子系统是最有效和最经济的调控在转录水平上的体现。该调控模型在生物学发展史上具有划时代意义,为基因表达调控这一难题的揭示奠定了基础。因而他们在1956年获得了诺贝尔奖。此外,他们还提出核糖体不含有负责氨基酸顺序装配的模板,认为每个DNA)顺反子引起寿命有限的RNA分子的合成,在此RNA的核苷酸序列中,包含着氨基酸序列的信息。随后该RNA与核糖体结合,给核糖体合成某一蛋白质的能力。由此进一步明确了基因的作用和遗传信息转移的中心法则,并由大量实验所证实。
(六)“跳跃基因”和“断裂基因”的发现 对于二倍体高等生物而言,能维持配子或配子体正常功能的最低数目的一套染色体就称为一个基因组,一个基因组中包含一整套基因。对于多数只有一条染色体的原核生物而言,它的整个染色体就是一个基因组。在20世纪50年代以前人们认为每一基因组的DNA是固定的,它包括数目固定、位置固定和功能固定的一系列基因,而且这些基因的位置和他们的功能无关。1961年大肠杆菌乳糖操纵子的发现就表明功能上相关的结构基因往往紧密排列在一起,说明大肠杆菌的染色体不是一个随机排列的集合体。50年代初,B.McClintock在玉米的控制因子的研究中已经指出某些遗传因子是可以转移位置的。60年代末在大肠杆菌中发现了可以转移位置的插入序列,接着在真核生物和原核生物中发现基因组中的某些成分位置的不固定性是一个普遍的现象,并将这些可转移位置的成分称为跳跃基因(jumping
gene),亦称转座因子(transposon element)。此外,传统的观点认为,一个结构基因是一段连续的DNA序列,70年代后期发现绝大多数真核生物基因都是不连续的,其中被一些不编码序列所隔开,故称为断裂基因。1978年,在噬菌体中还发现了重叠基因,一个基因序列可被包含在另一个基因中,两个基因序列可能部分重叠。
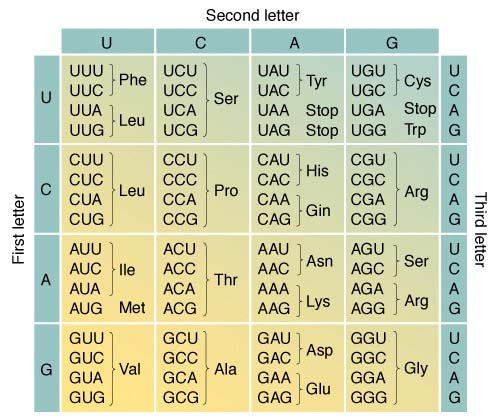
二.遗传密码
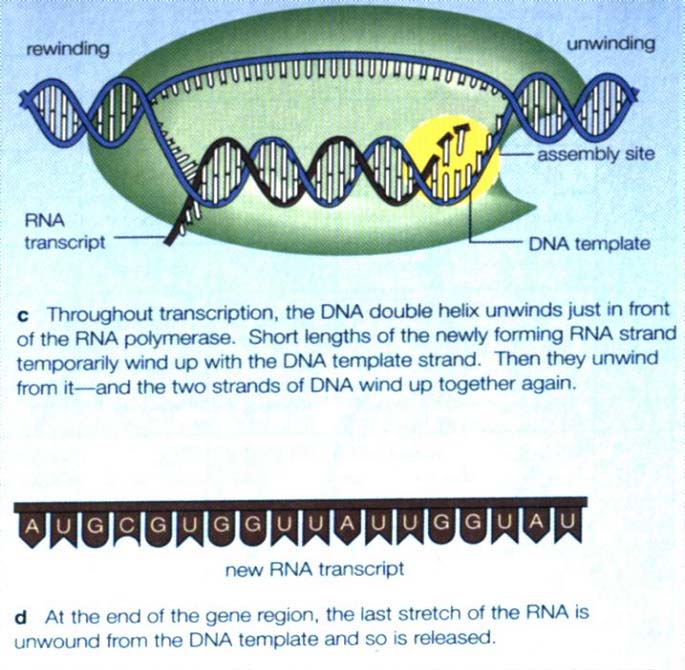
DNA中贮存的遗传信息先是转录给信使RNA(messenger RNA,mRNA),也就是以 DNA链为模板合成核苷酸序列与其互补的RNA分子,只是这里用尿嘧啶U代替胸腺嘧啶T,实现A―U配对。然后mRNA翻译成蛋白质。DNA中的遗传信息通过mRNA的介导而决定了蛋白质的一级结构。也就是说,DNA的核苷酸序列决定mRNA的核苷酸序列,mRNA的核苷酸序列决定蛋白质中的氨基酸序列。遗传密码的性质:
1.所有的遗传密码子都是由3个核苷酸组成的,所以可称三联体密码(triplet code);
2.许多氨基酸的密码子不止一个,因为4种核苷酸组成的密码子可以有43=64种,氨基酸却只有20种。几种密码子编码同一种氨基酸,这称为密码的简并性(degeneracy
of the code)。编码同一种氨基酸的两种以上的密码子称为简并密码子(degenerate
codon)。
3.64种密码子中除3种外都编码氨基酸。3种不编码氨基酸的密码子是UAA、UAG和UGA,它们的作用是终止mRNA翻译成蛋白质,所以称为终止密码子(stop
codon)。
4.密码子AUG除了编码甲硫氨酸(Met)外,还是mRNA翻译产生肽链时的起始位置。AUG是真核生物惟一的起始密码子(initiate
codon)。原核生物中,在很少的情况下,UGU也被用作起始密码子。
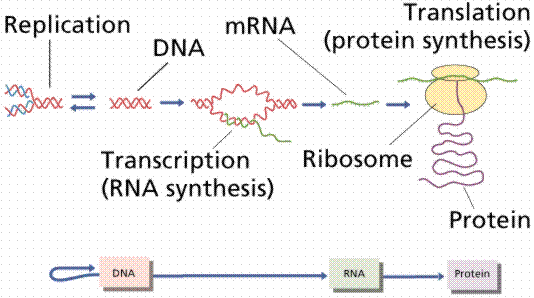
三.遗传信息的传递――复制和转录
DNA双链分子可以单链作为合成互补链的模板,从而把亲链中携带的遗传信息原样地传递给两条DNA子链。DNA双链分子可将携带的遗传信息通过互补的mRNA链传递给组成蛋白质分子的肽链。
因此,对于模板链(templete strand)有两种定义:
1.对于DNA复制而言,双链DNA中的两条链都是合成新的子链的模板链。
2.对于DNA转录而言,指导合成mRNA的那条DNA单链,也就是与mRNA互补的单链称为模板链或反义链(anti-sense
strand)。与 mRNA序列相同(除了T代替U)的那条DNA单链称为编码链(coding
strand)或有意义链(sense strand)。

四.中心法则
遗传物质可以是DNA,也可以是RNA。细胞的遗传物质都是DNA,只有一些病毒的遗传物质是RNA。这种以RNA为遗传物质的病毒称为反转录病毒(retrovirus),在这种病毒的感染周期中,单链的RNA分子在反转录酶(reverse
transcriptase)的作用下,可以反转录成单链的DNA,然后再以单链的DNA为模板生成双链DNA。双链DNA可以成为宿主细胞基因组的一部分,并同宿主细胞的基因组一起传递给子细胞。在反转录酶催化下,RNA分子产生与其序列互补的DNA分子,这种
DNA分子称为互补 DNA(complementary DNA),简写为 cDNA,这个过程即为反转录(reverse
transcription)。
遗传信息并不一定是从DNA单向地流向RNA,RNA携带的遗传信息同样也可以流向DNA。但是DNA和RNA中包含的遗传信息只是单向地流向蛋白质,迄今为止还没有发现蛋白质的信息逆向地流向核酸。这种遗传信息的流向,就是克里克概括的中心法则(central
dogma)的遗传学意义。